CAPÍTULO 2 ANATOMÍA Y FISIOLOGÍA J. Murube, Chen Zhuo |
| El aparato ocular se ha creado para
servir a la córnea. Consta de tres partes : (1) Un sistema de producción contituidos por
la tres clases de dacrioglándulas (acuoserosas, mucosas y lípidas). (2) un sistema de
trabajo, constituido por la cuenca lacrimal (córnea, saco conjuntival y párpados. (3) Un
sistema de emunción, constituido por las vías lacrimales (canalículos, saco lacrimal y
conducto lacrimonasal)
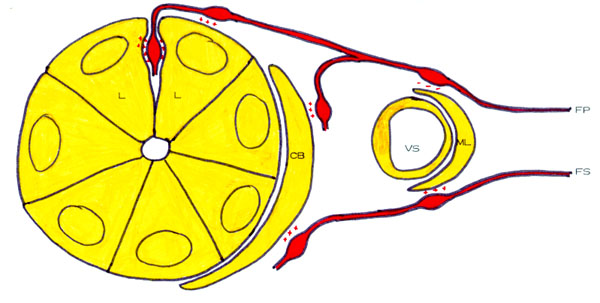
A. CUENCA LACRIMAL La cuenca lacrimal es el espacio comprendido entre los párpados y la superficie ocular, incluyendo la córnea y el saco conjuntival. Su cavidad está ocupada por un fluido de procedencia y componentes múltiples, el mar lacrimal (figura 2-1).
SUPERFICIE CORNEAL La córnea, especialmente su epitelio, es el centro del aparato lacrimal, para cuyo servicio ha surgido y evolucionado todo el complejo aparato lacrimal. La córnea tiene una superficie total de 110 mm2, es decir, el 6% de la superficie total de la superficie de la cuenca lacrimal. Un segmento superior de córnea de aproximadamente 1 mm de sagita está permanentemente cubierto por el párpado superior La córnea consta de 5 capas, que de delante a atrás son el epitelio, la membrana de Reichert-Bowman, el estroma, la membrana de Descemet y el endotelio, cuyos respectivos espesores son en el centro corneal de 60, 10, 450, 5 y 5 m m. De ellas, trataremos con más extensión la membrana de Reichert-Bowman y el epitelio. La membrada de Reichert-Bowman tiene un espesor de 0'04 40 m m en el centro corneal y 0'5 mm en la periferia. Es dura, resistente, y está perforada por numerosos puntos para permitir el paso de las terminaciones nerviosas. Una vez destruida, no se regenera. Sobre ella, se apoya el epitelio corneal. El epitelio corneal es estratificado escamoso no queratinizado. Está formado en profundidad por 5 o 6 células, de las que la más profunda constituye el estrato basal, las 2 ó 3 intermedias las células aladas, y las superficiales, el estrato apical, que contacta con el mar lacrimal. Las células basales miden 18 m m en sentido anteroposterior y 10 m m en sentido transversal. Su polo posterior es plano y está unido a la membrana basal por hemidesmosomas. Su polo anterior es convexo. Las paredes laterales son penta o exagonales, planas, y se unen a las células adyacentes por interdigitaciones, desmosomas y zónulas adherentes (gap junctions) y occludentes (tight junctions). Su núcleo es alargado paralelamente al eje mayor celular, y a veces tiene mitosis. En el citoplasma hay numerosos tonofilamentos de 8 nm de diámetro que, más o menos perpendiculares a la superficie corneal, terminan en los hemidesmosomas El estrato de células aladas tiene 2 o 3 células, aplastadas siguiendo la curvatura corneal; su cara anterior es convexa y su cara posterior tiene dos o mas concavidades para adaptarse al polo anterior de las células basales. La seción sagital histológica de las células les da el aspecto de una alas, de donde el nombre de Flügelzellen con que las bautizaron los histólogos alemanes. La membrana citoplásmica tiene muchas interdigitaciones y desmosomas. El eje del núcleo es paralelo a la superficie. El citoplasma posee abundantes tonofibrillas. El estrato superficial o de celulas apicales está formado por dos capas de células poligonales muy aplanadas, que miden unos 4 m m en sentido anteroposterior y 40 m m en el transversal. El núcleo es aplastado. El citoplasma tiene escasas organelas, abundantes tonofibrillas, abundantes depósitos de glucógeno y bastantes vesículas claras, algunas de ellas con contenido granular. La membrana citoplásmica de las células del estrato superficial se relaciona con las células vecinas por desmosomas y zónulas occludentes. La membrana anterior de las células más superficiales tiene abundantes microvilli y microplicae. Los microvilli tienen un diámetro de 0'10-0'15 m m y una longitud de 0'10-0'30 m m, y están separados unos de otros por 0'07-0'15 m m. Las microplicae están más dispersas y a veces se unen formando elevaciones anulares de 3-4 m m de diámetro. Las células superficiales del epitelio producen una glicoproteína glicosilada polianiónica, conocida como glycocalyx, que también se produce en la superficie de la conjuntiva y de otras mucosas (Gipson et al 1992). Cualquier alteración de las células epiteliales puede modificar esta mucina epitelial Células matrices. Las células matrices del epitelio corneal parecen estar localizadas en el anillo limbal que separa el epitelio corneal del conjuntival (figura 35-5). Esto tiene importancia en el ojo seco, y en general en la patología de la superficie ocular, y soporta las nacientes técnicas de transplantes limbales. Anatómicamente, el limbo es una zona de transición entre el epitelio corneal y conjuntival. Histológicamente este epitelio consta de más de 10 estratos el epitelio más grueso de la cuenca lacrimal- y su expresión fenotípica es un intermedio entre la de los epitelios de la córnea y la conjuntiva. Entre las células epiteliales hay abundantes melanocitos y células de Langerhans. El epitelio limbal carece de células caliciformes. Bajo el epitelio hay una rica red vascular, a partir de la que se supone le llegan los factores humorales reguladores. De antiguo había dos observaciones que necesitaban explicación: una era la migración de células epiteliales pigmentadas que forman en la córnea líneas que van desde el limbo hacia el centro; la otra, la reiterativa observación de Barraquer (1964, 1965, 1977, 1980) de que en el pterygion la conjuntiva es capaz de saltar sobre el limbo e invadir la córnea y de que esto debe tratarse con pequeños transplantes conjuntivales o corneoconjuntivales. Davanger et al sugirieron en 1979 que la estructura papilar limbal es la generadora de las células epiteliales corneales. Posteriormente, varios estudios soportaron la idea de que las células limbales emigran centrípetamente hacia la córnea para reparar pérdidas epiteliales traumáticas (Kinoshita et al 1981. Sobre estos hallazgos, Thoft et al 1983 elaboraron su hipótesis X,Y,Z para explicar cómo se genera y mantiene la masa epitelial corneal. Más tarde Buck 1985 observó que en el recambio epitelial normal hay también un movimiento celular centrípeto. En 1986 Schermer et al evidenciaron que las células matrices del epitelio corneal están en el limbo, basándose en la expresión de una queratina de 64 kd específica de la córnea. Actualmente, la suposición de que las células matrices del epitelio corneal están en el limbo se basa en: (1) las células basales del limbo son morfológica y bioquímicamente primitivas (Chung et al 1992), (2) las células limbales tienen un gran potencial de división (Cotsarelis et al 1987), (3) las células de renovación lenta LRC ("Label-retaining cells"), que sólo aparecen en los tejidos de renovación lenta, se encuentran en la capa basal del epitelio límbico, pero no en la del centro corneal (Cotsarelis et al 1989). Tseng y Zhang, 1995, encuentran que el 5-fluorouracilo es capaz de inhibir el crecimiento rápido de las células epiteliales del centro y periferia corneal, pero no el de las límbicas, (4) las células periférica se desplazan centrípetamente, apareciendo sus hemidesmosomas formando patrones radiales (Buck 1985), (5) las células de entre el limbo y la periferia corneal tienen un fenotipo similar al de las del limbo (Lauweryns et al 1983), (6) en la reparación de las heridas epiteliales corneales aparecen proliferación celular en el limbo adyacente (Kinoshita et al 1982, Murata et al 1993), (7) las células limbales autólogas, si se cultivan, reparan permanentemente el epitelio corneal dañado (Lindberg et al 1993) y (8) los transplantes limbales regeneran el epitelio corneal (Kenyon et al 1989). La razón por la que la naturaleza ha localizado en el área limbal a las células matrices del epitelio corneal puede ser (1) que allí tienen un rico soporte vascular, (2) que allí pueden ser muy pigmentadas, como de hecho lo son, para protegerse de las radiaciones actínicas (Lavker et al 1982), y (3) que allí actúan de barrera de protección cuando hay una herida corneal, para impedir que las células conjuntivales invadan la córnea para repararla (Chen et al 1991).
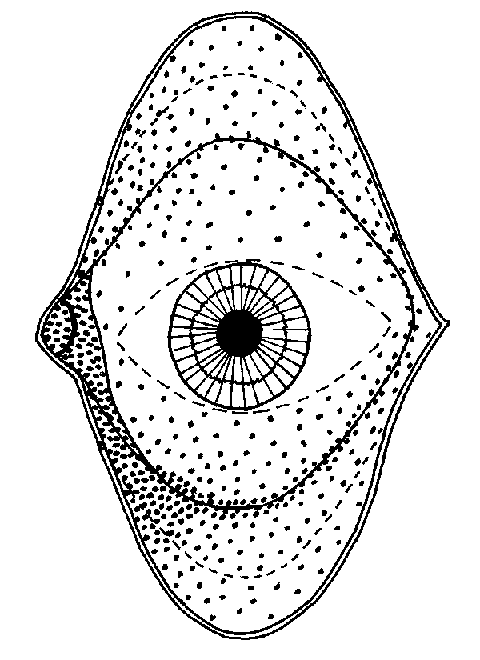
SUPERFICIE CONJUNTIVAL El saco conjuntival es la cavidad que queda entre los párpados y la superficie anterior del ojo con excepción de la córnea. Está tapizado por una mucosa: la conjuntiva o mucosa conjuntival. La estructura interior de la conjuntiva está formada por un corion y una epidermis. Es importante conocerla, puesto que las biopsias son pruebas frecuentes de diagnóstico en el ojo seco. El corion o dermis conjuntival tiene dos capas : Una profunda, gruesa, formada por tejido conjuntivo denso, por el que discurren vasos y nervios y que aloja o sirve de paso a las diversas glándulas de la conjuntiva. Otra más superficial, de cinco a diez veces más fina que la anterior y de un espesor igual al del epitelio. Su trama fundamental está formada por fibras colágenas y elásticas; posee abundantes capilares y sólo esboza papilas en las inmediaciones del limbo corneal. En su espesor hay linfocitos, histiocitos y plasmocitos. Allansmith et al 1976 consideran que en toda la conjuntiva hay un total de 2.100.000 plasmocitos, frente a los 3.380.000 que hay en las dacrioglándulas acuosas La epidermis conjuntival es un epitelio estratificado no queratinizado que asienta sobre una membrana basal que le separa del corion. Sobre ella hay un epitelio estratificado no queratinizado. Las células epidérmicas conjuntivales son algo distinta en la conjuntiva bulbar y en la palpebral. La epidermis de la conjuntiva bulbar es un epitelio pavimentoso estratificado de 5-8 capas, acentuándose la disposición pavimentosa en las inmediaciones del limbo corneal. Las células superficiales de la conjuntiva tienen forma hexagonal, de una diámetro medio de 8 m m, con numerosos microvilli. La epidermis de la conjuntiva palpebraly parte anterior de los fórnices es un epitelio mucoso cilíndrico de un espesor de dos células. El citoplasma de las células basales de la conjuntiva contiene melanina en el 20 % de los ojos secos y en el 10 % de los ojos normales (Álvarez et al 1986) se desconoce la finalidad de esta melanina, tal vez la de fotoprotección. La disposición topográfica en superficie de la conjuntiva muestra un área oculta por los párpados y un área expuesta al aire. El área oculta, o subpalpebral, es la que queda cubierta por los párpados cuando el globo ocular está en posición de ojo abierto y mirada al frente. La distancia media desde el limbo esclerocorneal al fondo de los fórnices es de 17'5 mm en el superior, 16'7 mm en el temporal, 14 mm en el inferior y 13'2 mm en el nasal (Pérez García, 1974). La superficie total de la conjuntiva es de 18 cm2. El fórnix superior, pese a ser muy móvil (al cerrar los ojos casi el 100% del cierre se hace a expensas de bajar el párpado superior), tiene escasos pliegues de flexión. Por el contrario, el fórnix inferior, pese a ser casi inmóvil, tiene muchos pliegues, lo que hizo sugerir (Murube 1981a) que la misión de los pliegues del fórnix inferior no es actuar como pliegues de flexión, sino aumentar la superficie de células caliciformes que producen mucina con finalidad limpiadora y dar al fórnix inferior la función de pozo de sedimentación y digestión del mar lacrimal. El área expuesta del ojo, -sólo oculta momentáneamente durante el parpadeo y continuadamente durante la connivencia-, está comprendida entre los párpados abiertos, y está formada por la córnea en el centro y dos triángulos o trígonos conjuntivales blancos uno a cada lado; la superficie del trígono lateral es más grande que la del medial. La superficie conjuntival total expuesta tienen un valor total de 0'8 cm2. Esta superficie, unida a la de la córnea expuesta, da un valor total a la superficie ocular expuesta de 1'8-2 cm2 en posición primaria de la mirada. En la mirada hacia arriba alcanza hasta 3 cm2 y en la mirada hacia abajo, 1 cm2. En cada trígono pueden aparecer, al lateralizar los globos oculares en su dirección, múltiples y pequeños pliegues conjuntivales verticales, a los que nunca se ha prestado atención, pero que pueden ser un importante signo para interpretar la participación de la conjuntiva en el ojo seco y otras patologías de la superficie ocular. A lo largo del margen palpebral aparecen otros pliegues, los pliegues labiformes, sobre los que se volverá a tratar en la parte dedicada al diagnóstico por fentobiomicroscopia. El lago lacrimal (lacus lacrimalis) es la porción de la cuenca lacrimal situada en el ángulo mediall de la hendidura palpebral, nasalmente a los puntos lacrimales. En él se encuentran, de nasal a temporal, la carúncula lacrimal y la plica semilunar. Expondremos con mayor extensión este área, pues en este libro hacemos la primera descripción de una nueva prueba diagnóstica para los síndromes de ojo seco : el borramiento de los surcos lacunares. La carúncula lacrimal (caruncula lacrimalis) (figura 2-2) es una excrecencia del tamaño de un grano de arroz, cuyo eje mayor, de 3'5 mm, está inclinado 45º hacia abajo y afuera. Parte de la carúncula está en área expuesta pero sus partes extremas superior e inferior son retropalpebrales, más en el extremo inferior. La parte posterior y medial de la carúncula es sésil y se continúa con los tejidos adyacentes. La parte superficial da hacia la cuenca lacrimal y está mojada por el mar lacrimal.
La carúncula tiene un núcleo de tejido conjuntivo fibroelástico denso, muy vascularizado, con algunas fibras musculares estriadas provenientes del músculo de Duverney-Horner. La parte sésil se continúa insensiblemente con el tejido adyacente. La parte nasal se separa del tejido cutáneo del canto medial por un surco en V horizontal. La parte lateral se separa de la plica lacrimal por un surco en Y vertical cuyas ramas inferior y lateral forman el surco carunculo-plical. La parte superficial de la carúncula, que da hacia la cuenca lacrimal, está cubierta por un tejido ectodérmico formado de un corion y un epitelio. El corion tiene abundantes linfocitos y plasmocitos. El epitelio, sustentado por una membrana basal, es de tipo plano estratificado no queratinizado y en la falda lateral de la carúncula va cambiando paulatinamente su estructura hasta tomar la del epitelio conjuntival, y hacerse cilíndrico biestratificado en el surco carunculo-plical. En el núcleo y corion caruncular hay 1-4 glándulas de Krause, glándulas (pseudo)sudoríparas de Moll, y aproximadamente 10 folículos pilosos, cada uno conteniendo 1-2 glándulas sebáceas, algunas de las cuales desembocan directamente en la superficie caruncular. Estas formaciones se exteriorizan a través del epitelio. En el epitelio también hay numerosas glándulas mucíparas superficiales o en criptas. Los linfáticos carunculares drenan a los ganglios submandibulares. La plica semilunar (plica semilunaris) es un repliegue falciforme de la conjuntiva bulbar. Su base, sésil, está situada inmediatamente por fuera de la carúncula, y su borde libre es cóncavo hacia el lado temporal y un poco hacia arriba. El borde libre de la plica mide aproximadamente 10 mm de longitud, de los que quedan 3 bajo el párpado superior, 2 en el área expuesta, y 5 bajo el párpado inferior. La dimensión horizontal de la plica en la zona expuesta es en la mirada de frente de unos 2 mm, haciéndose menor en la adducción y mayor en la abducción. El interior de la plica es una alcála de tejido mesodérmico submucoso muy escaso que contiene vasos linfáticos, vasos sanguíneos y nervios; ocasionalmente pueden haber una tenue lámina cartilaginosa y/o algunas fibras musculares, tal vez vestigios filogenéticos de cuando la plica funcionaba como membrana nictitante. Recubriendo las superficies de la plica está la mucosa conjuntival, con un corion mucoso y un epitelio sustentado por su membrana basal. El epitelio de la cara anterior de la plica es cilíndrico poliestratificado y posee numerosísimas células caliciformes, ya aisladas superficiales ya en criptas, especialmente en su mitad inferior. En la cuarta parte de las personas hay en el borde libre de la plica o en sus inmediaciones una cripta mucosa arboriforme que en ocasiones también tiene lacrimocitos de secreción acuoserosa. El epitelio de la cara posterior de la plica es cilíndrico biestratificado, como el de los fórnices conjuntivales y surco pericaruncular. La conjuntiva puede ser considerada como una glándula en sábana que vierte en la cuenca lacrimal la heterogénea secreción de sus múltiplas glándulas. Células matrices. Hay dos teorías sobre la localización de las células madres del epìtelio conjuntival: La teoría fornical sostiene que están localizadas en el fondo del fórnix conjuntival, desde donde sirven hacia delante la conjuntiva palpebral y hacia atrás la conjuntiva bulbar. Se basa esta teoría en que las células epiteliales de esta zona (1) tienen en el recién nacido gran actividad divisoria (Lavker et al 1991), (2) poseen más células LRC que ninguna otra de la conjuntiva (Tsai 1990), y tienen un potencial proliferativo muy superior al de las palpebrales y bulbares. La teoría margo-limbal sostiene que las células conjuntivales proceden de dos orígenes, uno situado en la junción mucocutánea del borde palpebral y otra en el área limbal pericorneal. Las células que se generan aquí se desplazan y mueren en el limbo (Perér et al 1996).
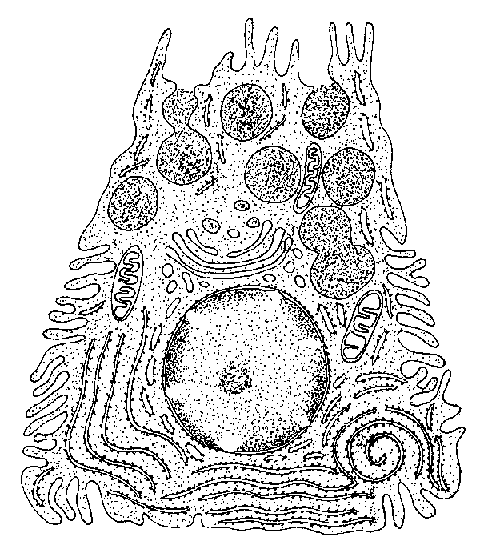
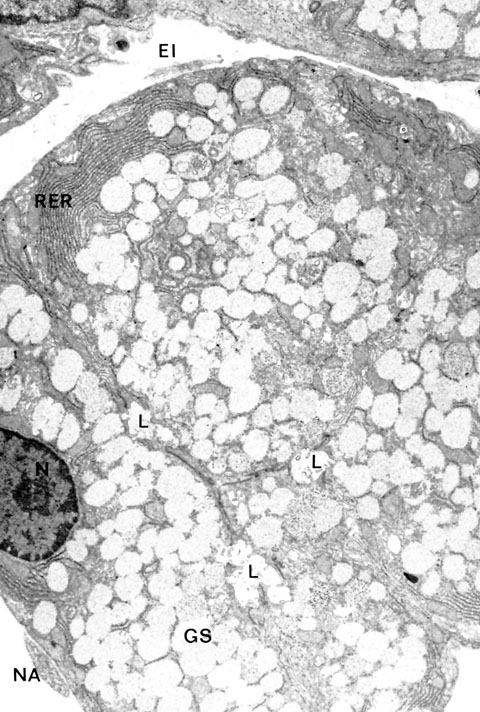
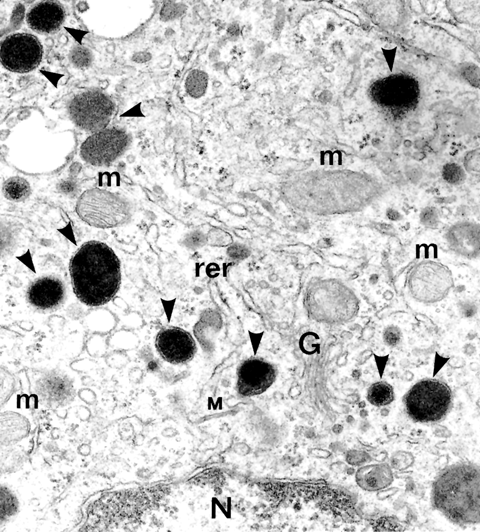
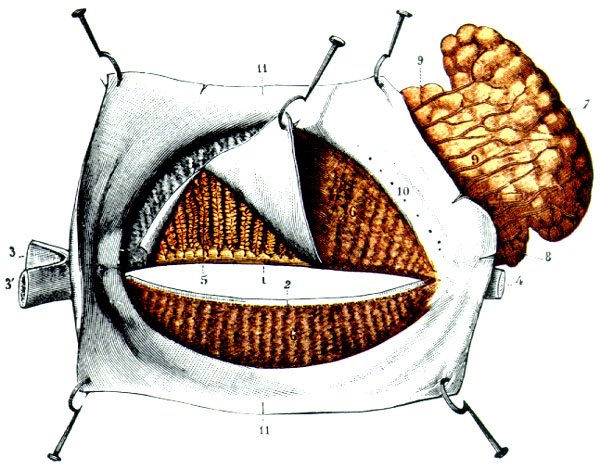
B. DACRIOGLÁNDULAS ACUOSAS DISTRIBUCIÓN Y ANATOMÍA Las glándulas lacrimales acuosas, o mejor, acuoserosas, constan de una glándula lacrimal principal y varias glándulas lacrimales accesorias. La glándula lacrimal principal está alojada en la parte anterior del ángulo supero-lateral de la órbita ósea. Es una glándula arracimada tubuloacinosa con una porción palpebral de unos 0'6 ml de volumen y una porción palpebral de unos 0'45 ml de volumen. La glándula tiene un parénquima secretor exocrino y un estroma de servicio. El parénquima exocrino está compuesto por celulas secretoras o lacrimocitos (figura 2-3). Un lacrimocito tiene forma de pirámide truncada, y mide 4 m m de ancho por 10 m m de alto. Su membrana citoplásmica tiene una cara basal unida a la membrana basal del acino, y ocasionalmente a células de Boll, por hemidesmosomas; las caras laterales, que son de 4 a 6, contactan con lacrimocitos vecinos, con los que se relacionan por interdigitaciones, desmosomas y zónulas occludentes y adherentes; la cara luminal presenta expansiones citoplásmicas digitiformes (microvilli). El núcleo está cerca de la cara basal. El citoplasma contiene un citoesqueleto; el citoesqueleto externo, como el de las demás células acinares cromafines y la mayoría de las células eucarióticas, contiene abundantemente una proteína, la fodrina, cuya subunidad a -fodrina posee sitios de unión para actina, calmodulina y CD45; la a -fodrina interviene importantemente en el proceso secretorio del lacrimocito y posiblemente en la aparición y evolución del síndrome de Sjögren. En el interior del citoplasma se encuentra el retículo endoplásmico -casi todo él de tipo rugoso-, aparato de Golgi, mitocondrias, lisosomas y gránulos secretorios.
Los lacrimocitos se disponen en una capa monoestratificada, formando saquitos secretores o acinos (figura 2-4). Cada acino está rodeado por una membrana basal. Entre la superficie basal de los lacrimocitos y la membrana basal se interponen espaciadamente células mioepiteliales de Boll, que tienen capacidad contráctil. Entre las células secretorias exocrinas, nosotros describimos en 1982 la presencia de células secretorias endocrinas pertenecientes al sistema APUD (figura 2-5) (Murube 1982, pp 344-345). Varios acinos con sus estroma y sus dúctulos constituyen un lobulillo glandular. Varios lobulillos, delimitados por una membrana propia, constituyen un lóbulo, y varios lóbulos, forman la glándula
Las superficies secretoras de los lacrimocitos delimitan el lumen o túbulo acinar. Los túbulos de diversos acinos desembocan en un dúctulo, cuyo epitelio sigue siendo secretor; varios dúctulos se reúnen formando dacriodocos, que corren por el espesor de los tabiques interlobares y desembocan en la cuenca lacrimal. Las glándulas lacrimales accesorias son casi medio centenar de pequeños acúmulos glandulares. Las distribuidas a todo lo ancho de los fórnices conjuntivales (llamadas glándulas de Krause) son unas 25 en el fórnix superior, 4 en el inferior, y 1 en la carúncula. Las distribuidas en los párpados (llamadas glándulas de Wolfring-Ciaccio) son unas 10 en el párpado superior y unas 3 en el inferior. Los números citados son medias y hay grandes variaciones numéricas individuales. El volúmen total de todas las glándulas accesorias es de 0'1 ml. La morfología histológica es parecida a la de las glándulas lacrimales principales, siendo las diferencias más notorias hasta el presente descubiertas la mayor abundancia de células plasmáticas en la glándula principal (3.200.000 plasmocélulas en la glándula principal y sólo 180.000 en todas las accesorias, según Allansmith et al 1966).
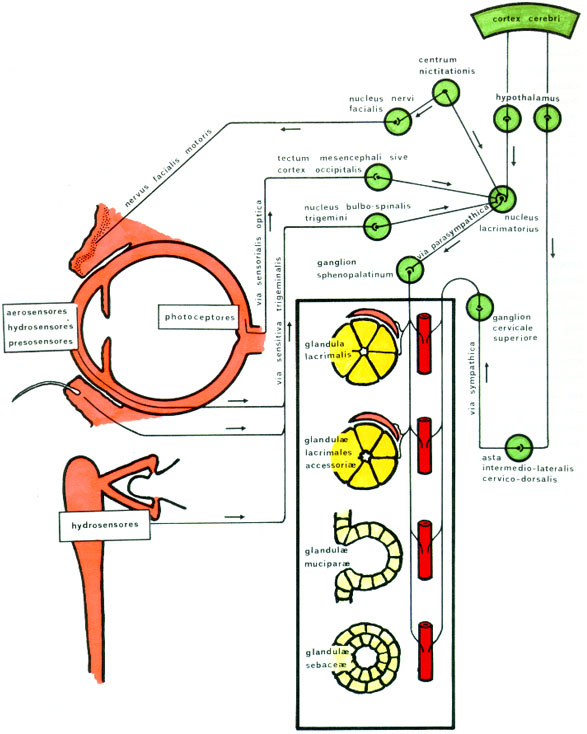
CONTROL NERVIOSO DE LA SECRECIÓN LACRIMAL Inervación de las glándulas lacrimales No está bien probado si las glándulas lacrimales acesorias no tienen inervación secretoria, por lo que sólo producirían secreción basal, mientras que la glándula principal, con inervación secretoria, produce secreciones basal, refleja y emocional. Incluso desde el punto de vista funcional no se está seguro si la porción palpebral de la llamada glándula principal no es sino un acúmulo de lobulillos de glándulas accesorias que sólo actúa en la secreción basal Los neurotrasmisores liberados en las terminaciones nerviosas autonómicas de las glándulas lacrimales (figura 2-6) son colinérgicos, adrenérgicos y peptidérgicos
Las terminaciones colinérgicas o parasimpáticas están en las caras basal e intercelulares de los lacrimocitos, en las celulas mioepiteliales de Boll y en las células musculares lisas de las arteriolas de la glándula. Cuando se estimulan, producen simultaneamente secreción de los lacrimocitos, expresión de los acinos por contracción de la células de Boll, y vasodilatación de las arteriolas lacrimales. Las terminaciones adrenérgicas se sabe que existen en las glándulas lacrimales, pero es discutido si se encuentran en los lacrimocitos. Nosotros (Murube 1981a, p 189) no hemos encontrado terminaciones simpáticas en los lacrimocitos. Las terminaciones peptidérgicas responden a ciertos péptidos, de los que el más conocido es el VIP (vasoactive intestinal polypeptid), un potente estimulador de la secreción acuosa y proteica de la glándula lacrimal. Actúa aumentando el cAMP intracelular y activando la proteinkinasa cAMP-dependiente (PKA). Los VIP-receptores tipo I y II aparecen en la rata en las membranas basal y laterales de los lacrimocitos, y en las células ductales. Los VIP-receptores tipo II son los únicos que se encuentran en las células mioepiteliales de los acini (Hodgeset al 1996). Otros péptidos detectados en las glándulas lacrimales son la substancia P, el CGRP (calcitonin gene-related peptide) y la 5-hidroxitriptamina (que es un mediador de los mastocitos). También hay alguna evidencia de que existe NPY (neuropéptido Y). Todos ellos están muy relacionados con el metabolismo de los mastocitos. Por ello, el cromoglicato sódico, que es un estabilizador de los mastocitos, reduce la respuesta secretoria peptidérgica.
Centros superiores de la lacrimación (figura 2-7) El complejo mecanismo de la lacrimación emocional probablemente requiere conexiones multicéntricas. Se han detectado áreas relacionadas con la lacrimación emocional en el córtex límbico, en el córtex frontal, en el tálamo óptico y en la región hipotalámica. De allí saldrían las fibras que conectarían con los núcleos y vías lacrimosecretoras parasimpática y simpática
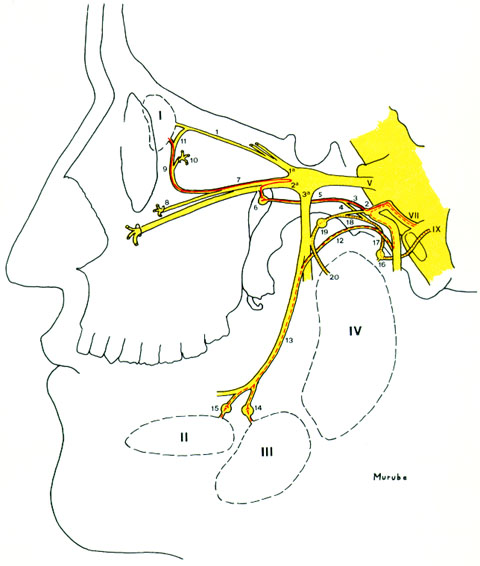
Vía parasimpática (figuras 2-8 y 12-1) El núcleo parasimpático preganglionar lacrimosecretor está situado en la parte ventral del núcleo salival superior, en el rombencéfalo. De sus cuerpos neuronales salen unos axones de 9-10 cm de longitud, que siguiendo la vía del facial, se separan de él a nivel del ganglio geniculado en el interior del peñasco, siguen por el nervio petroso superficial mayor, continúan por el nervio vidiano y finalmente llegan al ganglio esfenopalatino situado en la fosa pterigopalatina que separa el macizo facial del craneal. Allí, sinapsan con los cuerpos neuronales de la segunda neurona. De ellos salen los axones postgangliónicos, de unos 5 cm de longitud, que saltan a la segunda rama del trigémino y haciendo o sin hacer un trayecto de ida y vuelta que pasa por el ganglio de Gasser, siguen por la segunda rama del trigémito, y penetran en la órbita por la rama orbitocigomática, continúan por su ramo anastomótico al nervio lacrimal (rama de la primera rama del trigémino), y directamente o a través del nervio lacrimal alcanzan la glándula lacrimal principal.
Vía simpática (figura 2-9) El núcleo simpático preganglionar está situado en la columna gris del asta intermedio lateral de la médula cervicodorsal, escalonado entre la D-I y la D-IV. Sus axones siguen por los ramicomunicantes blancos D1, D2 y D3 , ascienden por la cadena simpática paravertebral y alcanzan el ganglio cervical superior, donde sinapsan con los cuerpos neuronales de la segunda neurona. De ellos salen los axones postgangliónicos que saltan al plexo pericarotídeo. Desde allí alcanzan la glándula lacrimal y los vasos de la cuenca lacrimal por la arteria oftálmica y sus rama, por anastomosis con el ganglio de Gasser y primera rama del trigémino, y por anastomosis con el nervio vidiano y vía parasimpática.
MECANISMO DE LA SECRECIÓN LACRIMAL El líquido lacrimal secregado en las glándulas acuoserosas está compuesto mayoritariamente por agua, y en menor cantidad por proteínas, sales, glucosa, etc. Estos componentes no se vierten al lumen intraglandular por un proceso similar para todos ellos, sino que lo hacen por algunos de los tres siguientes mecanismos: (1) Hidroiónico. La acetilcolina liberada en las terminaciones efectoras muscarínicas del parasimpático lacrimosecretor es captada por la membrana de los lacrimocitos, lo qe altera su permeabilidad iónica y aparecen los cambios de potencial de membrana conocidos como potenciales de secreción. Como resultado global, las células eliminan a los túbulos potasio y cloro, y con ellos agua. (2) Exocítico. Los lacrimocitos sintetizan las proteínas específicas de la lágrima en su retículo endoplásmico rugoso; este material pasa a las vesículas del aparato de Golgi, donde se unen a glúcidos para formar glucoproteínas y mucopolisacáridos. Del aparato de Golgi se desprenden microvesículas que progresan por el citoplasma, al tiempo que se unen, coalescen y forman gránulos secretorios mayores cuyo contenido va madurando. Finalmente, la membrana de los glánulos secretorios se funde a la membrana citoplásmica luminal y su contenido se vierte al lúmen del acino. Así se secretan las mucinas de la dacrioglándulas acuoserosas, las lipocalinas, la lisozima, la pieza o componente secretorio que liga los dímeros de IgA, etc. (3) Osmótico. El líquido vertido a los lúmina glandulares por los mecanismos hidroiónico y exocítico es fuertemente hipertónico con respecto al suero. Así, aparece una corriente acuosa a través del epitelio tubular, ductular y de los dacriodocos, de forma que cuando la lágrima alcanza la desembocadura de los dacriodocos en la cuenca lacrimal es prácticamente isotónica con el suero. El líquido intercelular, al ser atraído hacia los conductos secretores intraglandulares, arrastra solutos no iónicos, entre los que destaca la glucosa y algunos enzimas. No se sabe mucho sobre las diferencias de secreción de las glándulas lacrimales principales y accesorias. Se sabe que ambas producen la pieza secretoria de la IgA-s.
CANTIDAD DE SECRECIÓN La producción lacrimal acuoserosa por minuto en circunstancias normales basales durante la vigilia se ha calculado muy diversamente según los autores : 0'6-1'1 m l (Saracco et al 1974), 0'5-2'2 m l (Göbbels et al 1992), 1'2 m l (Ehlers 1967, Ehlers et al 1972), 5-10 m l (Murube 1981a, p 406), 10 mg (Nover et al 1952, Norn 1965), etc. Suponiendo un volumen de 5-10 m l/min, en 16 horas de vigilia la cantidad de producción acuoserosa asciende a 5-10 ml. Durante el sueño el ojo no segrega lágrima (Schirmer 1903), o lo hace en cantidades mínimas (Szmyt 1958). Esta escasa secreción nocturna no se hace de una manera uniformemente constante, sino intermitentemente (Murube 1981a, p 408). Las variaciones entre sexos son contradictorias, inexistentes o estadísticamente no valorables (Henderson et al 1950). La secreción basal tiene claras variaciones etarias. Se inicia ya durante los últimos meses del embarazo, y los niños prematuros viables la tienen, aunque menor que los nacidos a término (Kästner 1957). El máximo de secreción lacrimal se tiene en las 3 primeras décadas de la vida. Y después disminuye paulatinamente. En la siguiente tabla se ven los resultados en 8 grupos etarios, por décadas, de 20 individuos normales en cada grupo, con tiras de papel de filtro de 50+5 mm.
VARIEDADES DE LACRIMACIÓN Se conocen 3 clases de lacrimación : la basal, la refleja y la emocional. La lacrimación basal es la que el ojo produce en circunstancias basales. Tiene muchas variedades fisiológicas (con ojos abiertos, con ojos cerrados, durante el sueño) y no fisiológicas (tras instilar anestesia, tras denervación pregangliónica, postgangliónica o sensorial). La secreción lacrimal basal se inicia durante los últimos meses del embarazo, estando presente en los fetos prematuros viables (Kästner 1957). La lacrimación refleja es la estimulada a través de un reflejo nervioso nacido en la superficie ocular y en la retina, pero también en fosa nasal (Schirmer 1903), tracto respiratorio (Gaynon 1968), boca (Allansmith et al 1971), faringe (Demtschenko 1872), y ocasionalmente en áreas somáticas de nervios periféricos como el reflejo auro-lacrimal (Arboleda 1914) o el reflejo cloaco-lacrimal (Murube 1980). Nosotros describimos el reflejo oculo-lacrimal (Murube1977a), que se produce al comprimir un ojo; como la secreción lacrimal que produce es bilateral, lo utilizamos para sacar lágrima (para analizarla) de un ojo en los pacientes de ojo seco, comprimiento el ojo cerrado contralateral. La secreción lacrimal refleja se inicia a los 15-30 días de nacer, con grandes variaciones individuales, pues puede haber neonatos que ya la tienen o niños de más de un mes, en los que aún no existe. La lacrimación emocional es la que se produce a través de una orden cerebral relacionada con determinados estados anímicos. La lacrimación emocional es filogenéticamente muy reciente, y sólo existe en la especie humana. Hay 5 teorías para explicar su aparición, todas basadas en la transformación del lagrimeo en un reflejo condicionado asociado a la angustia del llanto: La compresión de las glándulas lacrimales al comprimir las glándulas con el blefarospasmo del llanto (Darwin 1872), la liberación catártica de un estímulo nervioso por vía parasimpático-lacrimal (Freud 1893, 1929), la humidificación del aparato fonatorio (Montagu 1969), el aclaramiento de productos biológicos liberados a la sangre por la emoción (Frey et al 1983), y la simbolización de la lágrima como manifestación de dolor, derivada de que el derramamiento de lágrima por lagrimeo reflejo se hace siempre asociado a dolor (Murube). Más tarde, la lágrima emocional, que inicialmente se exhibía sólo en situaciones de petición de ayuda y dolor, sirvió también para expresar lo contrario, es decir, el ofrecimiento de ayuda y solidaridad (Murube 1990a) la inversión de un símbolo facial para expresar lo contrario también se ha dado en otras exhibiciones faciales, como el enseñar los dientes, que asociado inicialmente al ataque, sirvió más tarde para expresar también con la sonrisa el beneplácito y la simpatía. La secreción emocional se admitía hasta el presente que se inicia a las pocas semanas o meses del nacimiento. Desde que se descompuso la lacrimal emocional en dos tipos, el de petición de ayuda y el de donación de ayuda (Murube et al 1990a), esta fecha queda asignada a la lacrimación de petición de ayuda, apareciendo la de donación de ayuda al acceso del uso de razón, es decir, entre los 5 y 7 años.
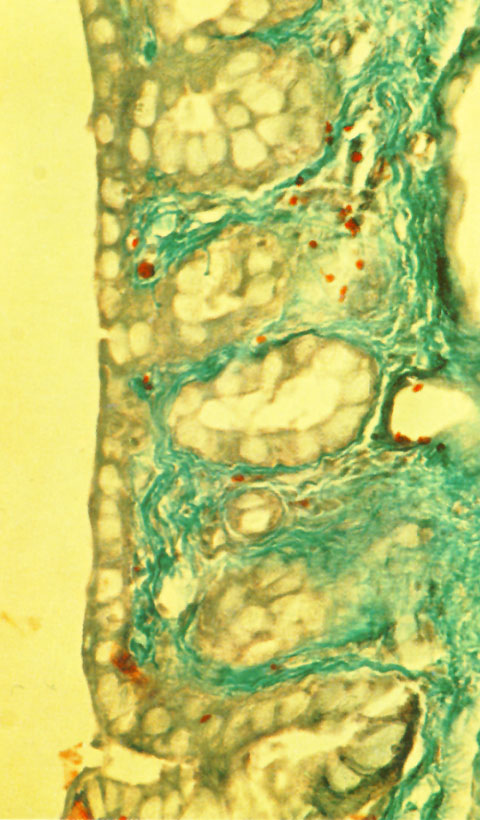
C. DACRIOGLÁNDULAS MUCOSAS DISTRIBUCIÓN Las dacrioglándulas mucosas están constituidas en cada ojo por millón y medio de células epiteliales mucíparas, llamadas células caliciformes. Las células caliciformes se disponen en la superficie conjuntival o en pequeñas oquedades del epitelio conjuntival, llamadas criptas mucosas de Henle (figura 2-10).
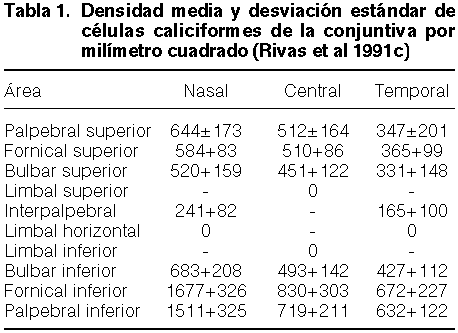
La células caliciformes están distribuidas con densidad diversa por toda la superficie conjuntival, pero faltan o son muy escasa en el anillo conjuntival perilímbico. Se distribuyen cuantitivamente según la tabla 1 (figura 2-11).
Según se ve, la máxima concentración de células caliciformes está en la parte medial del fórnix y párpado inferiores. El área de máxima concentración de células caliciformes es, para Green 1894, la conjuntiva de los fórnices y los tarsos para Wolff 1954, el fórnix inferior y la plica semilunar para Podhorányi 1966, los fórnices; y para Kessing 1968, la plica semilunar y la parte medial del fornix y tarso inferiores. A la vista de esta distribución, creemos que debe reconsiderarse la función principal de la mucina lacrimal. Si su función principal fuese el bajar la tensión superficial de la película lacrimal y el distribuirse sobre la córnea, su mayor concentración estaría aguas arriba de la córnea, en la región subpalpebral central superior y lateral superior. Su máxima concentración está si embargo aguas abajo de la córnea; al menos en este lugar, su función principal podría ser englobadora de cuerpos extraños, antimicrobiana y degradadora de los componentes lacrimales. En tal sentido, el fórnix inferior y el lago lacrimal sería como el pozo negro del mar lacrimal.
CICLO SECRETOR DE LAS CÉLULAS MUCÍPARAS Las células caliciformes se forman por mitosis a partir de células madres situadas en el estrato basal del epitelio conjuntival. Empujadas por la formación de nuevas células, se desplazan hacia la superficie, y a lo largo de este camino van madurando. Llegadas a la superficie liberan durante unos pocos días su secreción, y finalmente se desprenden a la cuenca lacrimal y son eliminadas con la lágrima Al reproducirse, la célula mucípara carece de mucina. En una segunda fase, cuando la célula aún no ha alcanzado la superficie, comienza a formar mucina: el núcleo se rechaza hacia la base, el aparato de Golgi produce y desprende gránulos secretorios; como consecuencia, la célula aumenta de volumen. En una tercera fase la célula , ya llegada a la superficie abierta o de una cripta mucosa de Henle, vierte su contenido al mar lacrimal por un mecanismo merocrino. Esta actividad se mantiene durante unos días. En una cuarta fase, la célula deja de sintetizar y segregar mucina, y se descama a la cuenca lacrimal. Las células caliciformes son muy sensibles a la vitamina A. La falta de vitamina A altera la secreción de mucina y degenera las células caliciformes.
VARIEDADES DE MUCUS CONJUNTIVAL El mucus conjuntival está fundamentalmente formado por glucoproteínas. Las glucoproteínas, como su nombre indica, tienen un componente proteico y otro hidrocarbonado. El componente proteico, que constituye algo menos del 50% de la glucoproteína, es una larga cadena polipeptídica, cuyos aminoácidos más abundantes son serina, treonina y prolina, y en la que también participan residuos de cisteína, cuyos grupos sulfhidrilos forman puentes disulfuro con grupos similares. Estas cadenas polipeptídicas están ligadas entre sí por sus N y C terminales y por interacciones entre las cadenas hidrofóbicas de sus aminoácidos. El componente azucarado, que constituye algo más de la mitad de la glucoproteína, forma alrededor de la cadena polipeptídica una cubierta incompleta de cortas cadenas laterales de carbohidratos, ligadas a la cadena polipeptídica por enlaces covalentes. Entre estos oligosacáridos están el ácido siálico, fucosa, galactosa, N-acetilglucosamina y N-acetilgalactosamina. El grupo extremo de estas cadenas laterales suele ser un ácido siálico. El ácido N-acetilneuramínico, al pH lacrimal, está completamente ionizado, aportando a las moléculas una densa carga negativa y una clara asimetría axial. El componente azucarado de las mucinas varía según la misión que estas mucinas hayan de cumplir. En el cuerpo humano se han aislado 9 genes diferentes para mucinas (MUC 1, MUC 2, MUC3, MUC4, MUC5AC, MUC5B, MUC6, MUC7 y MUC8). Estas mucinas son glucoproteínas altamente (hasta 50-60%) O-glicosiladas, y de ellas, una (MUC1) es una mucina no gelificante que recubre la membrana celular; cinco (MUC- 2, 3, 5AC, 5B y 6) tienen un alto contenido en cisteína apto para formar enlaces disulfuro y formar geles; y dos (MUC 4 y 8) son poco conocidas en cuanto a su estructura y funcionamiento Se sabe (Gipson et al 1996) que el epitelio corneal y conjuntival forma MUC1, que el epitelio conjuntival expresa MUC4, y que las células caliciformes conjuntivales expresan MUC5AC. La MUC1, de acuerdo a Sack et al 1996, es similar a la encontrada en la saliva, y se localiza en la membrana citoplásmica de las células epiteliales aumentando en abundancia desde el polo basal al apical de la célula. Esta mucina disminuye la capacidad de los microorganismos para adherirse al epitelio y es la última línea de defensa contra las proteasas de estos microorganismos Las mucinas de la lágrima de animales con KCS es diferente de la de los normales. Así, las cadenas oligosacáridas laterales de perros con KCS son más largas, tienen mayor proporción de fucosa, N-acetilglucosamina y galactosa en relación con la N-acetilgalactosamina, y una mayor cantidad de serina (Carrington et al 1996). Esta diferencia estructural puede justificar que la mucina de la KCS proporcione un menor efecto lubricante y de barrera.
ESTIMULACIÓN SECRETORA Las células mucíparas cerecen de terminaciones nerviosas secretoras. Es posible que la lágrima acuoserosa producida en las glándulas lacrimales contenga una macromolécula (probablemente un polipéptido) que, puesta en contacto con las células mucíparas a través del mar lacrimal, estimule la producción de mucina. El carbachol es un agente parasimpatomimético que estimula la secreción acuoserosa, y que por un mecanismo no aclarado, estimula también la secreción mucínica. La vasodilatación de la conjuntiva tiene un efecto positivo sobre la secreción de mucina. También lo tienen muchas situación no homeostática de las células caliciformes : alteraciones tisulares intersticiales, alteraciones del líquido lacrimal e irritaciones mecánicas. La mayor producción de mucina durante el día que durante la noche es posible que esté en relacion con esto, con factores humorales o con la acción mecánica del parpadeo.
FUNCIÓN DE LA MUCINA La función de la mucina parece ser múltiple: (1) Formar una capa protectora húmeda sobre el epitelio, debido a su capacidad de ligarse al agua y de retenerla. (2) Bajar la tensión superficial del componente acuoso de la lágrima, y favorecer que ésta se extienda sobre la superficie corneal y conjuntival. (3) Lubricar la córnea para facilitar el deslizamiento palebral. (4) Atrapar microorganismos y cuerpos extraños y arrastrarlos hacia el lago lacrimal. (5) Degradar algunos componentes lacrimales, especialmente los lípidos. En pH algo ácido, la mucina suele estar en forma de gel, y en pH alcalino, de sol. En forma de gel, la mucina es mejor protector y limpiador, y en forma de sol, mejor lubricante.
CANTIDAD DE MUCINA CONJUNTIVAL La población de células caliciformes de un ojo es de 1'5 millones, y su volumen global, de 2'2 m l. Como una célula caliciforme segrega al día una cantidad de mucina aproximadamente igual a su volumen, se supone que la secreción de mucina de cada ojo es aproximadamente de 2-3 m l/día (Ehlers et al 1972).
D. DACRIOGLÁNDULAS LÍPIDAS Están situadas en las aristas posterior (glándulas de Meibomio) y anterior (glándulas de Zeis y de Moll) del margen palpebral, y en la carúncula (glándulas de Zeis, glándulas de Moll).
DACRIOGLÁNDULAS LÍPIDAS POSTERIORES Las dacrioglándulas lípidas posteriores o glándulas de Meibomio (figura 2-12) están situadas en la parte porterior del margen palpebral. Son glándulas tubulo-acinares de secreción sebácea, encamadas en los tarsos, que vierten su contenido en el borde palpebral. Su número, según Sappey 1853, es de unas 30 en el párpado superior y unas 25 en el inferior.Tienen 5-10 mm de longitud por 0'3 de anchura , y el final de su túbulo excretor tiene 0'1 mm de diámetro, desembocando en el borde posterior del borde libre. Las bocas, separadas entre sí aproximadamente por 1 mm, aparecen 0'25 mm por delante de la arista posterior del borde libre. El epitelio por delante de las bocas es de tipo queratinizado, y por detrás no queratinizado.
Su mecanismo de secreción es holocrino, es decir, la célula se llena de producto secretorio, y después muere y se desprende hacia el exterior, donde paulatinamente se desintegra. La composición normal de los lípidos meibomianos es ésteres céricos 68%, ésteres de colesterol 16%, triglicéridos 6%, fosfolípìdos 4%, esfingolípidos 2%, y trazas de otros componentes. El segregado se vierte sobre le borde palpebral y forma la capa lípida de la película lacrimal. Las glándulas de Meibomio secretan por estímulos hormonales (tiene receptores androgénicos), neurales (tiene influjos colinérgico y VIP-érgico) y físicos (parpadeo, frote). Tienen una rica inervación sensitiva que incluye fibras CGRP (calcitonin gene-related peptid) y SP (substance P). Parece que están desconectadas del sistema neural reflejo, por lo que carecen de secreción refleja. Los vasos sanguíneos vecinos tienen inervación simpática NPY-érgica y TH-érgica. Las glándulas de Meibomio carecen de fibras contráctiles, por lo que liberan su contenido por la vis a tergo y por la compresión del parpadeo; por ello, su secreción se acumula durante el sueño en el lumen glandular, y se libera al despertar y reiniciar el parpadeo. Con la edad, la mayor flaccidez de los párpados y la menor fuerza del papadeo dificulta la salida de la grasa meibomiana
DACRIOGLÁNDULAS LÍPIDAS ANTERIORES Las dacrioglándulas lípidas anteriores son las glándulas de Zeis y de Moll. Tal vez participan en escasa cuantía en la capa lípida de la película lacrimal, llegando a ella tanto por difusión hacia atrás por el borde libre del párpado, como por avanzamiento del menisco lacrimal durante la fase de cierre del parpadeo. Las glándulas de Zeis son glándulas acinoalveolares que desembocan en el interior de los folículos de las pestañas y de la carúncula, generalmente dos por folículo. Algunas pocas desembocan directamente en el borde palpebral. Su número total es aproximadamente de 250. Segregan por un mecanismo holocrino Las glándulas de Zeis se cree no tienen inervación secretoria. La administración de andrógenos aumenta claramente su producción de sebo y el tamaño de las glándulas, mientras que los estrógenos reducen ambas características Las glándulas de Moll son glándulas tubuloalveolares apocrinas arrolladas en ovillo. Esta morfología hizo que se las llamase erróneamente glándulas sudoríparas apocrinas no deben ser confundidas con las verdaderas glándulas sudoríparas, que son eccrinas, y cuya misión es la termorregulación. Las glándulas de Moll están situadas en la parte anterior del borde palebral. Unas desembocan en los folículos de las pestañas y otras directamente en el borde libre de los párpados. También las hay en la carúncula lacrimal. Segregan por mecanismo apocrino (desprendiendo en el lumen glandular su polo apical repleto de segregado; este concepto está hoy en revisión). Su producción secretoria tienen un importante componente lípido, por lo que aunque no es el único, se las clasifica como dacrioglándulas lípidas Se desconoce su función principal. Tal vez fue feromónica (Murube 1981 p 354, 1987). Son similares a las glándulas pseudosudoríparas apocrinas de carúncula lacrimal, axila, periné, ingle, monte de Venus, prepucio, escroto, pezón y areola mamaria, y como ellas, comienzan a funcionar en la pubertad, lo que apoya su función feromónica. A este respecto, es posible que las largas pestañas como signo de feminidad expresen indirectamente la capacidad reproductora de la hembra fértil, y que el guiño procaz de un ojo sea un inconsciente símbolo de la expresión de estas glándulas y consiguiente liberación feromónica. Se desconoce si las glándulas de Moll tienen inervación secretoria. La mayoría de las glándulas sudoríparas apocrinas (axila, prepucio, pezón) tienen inervación simpática, pero algunas (axila) también la tienen parasimpática.
E. MAR LACRIMAL GENERALIDADES El mar lacrimal es el contenido líquido de la cuenca lacrimal, formado mayoritariamente por el conjunto de todas las secreciones de las dacrioglándulas acuoserosas, mucosas y lípidas. A ello hay que añadir un componente extraglandular minoritario que le llega a través de las paredes de la cuenca lacrimal : Ya Janin 1772 supuso que parte del líquido que llena la cuenca lacrimal no procede de las glándulas, sino de las paredes de la conjuntiva y de la córnea, Michel 1884 señaló que la continua evaporación del mar lacrimal aumentaría su osmolaridad y atraería liquido de los tejidos vecinos, Schirmer 1903 sugirió que la conjuntiva normal segrega en cantidad suficiente para humedecerse a sí misma, Charlton 1920 indicó que parte del humor acuoso pasa al mar lacrimal, y Ridley (1930) calculó que esto sería en cantidad de 0'81 m l/min cuando la presión intraocular es de 25 mmHg. El volumen del mar lacrimal es, según Fehér 1993, de 5-10 m l, de los que 4'5 m l son subpalpebrales, 2'9 m l están en los meniscos y 1'1 m l en la película lacrimal precorneal. Según Göbbels et al 1992, el volumen del mar lacrimal es de 4-13 ml.
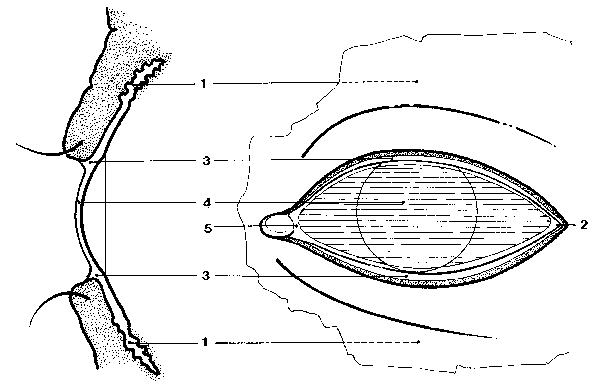
MORFOLOGÍA DEL MAR LACRIMAL El mar lacrimal (figura 2-1) tiene una zona oculta bajo los párpados que es el mar sub o retropalpebral y otra expuesta, que es el mar expuesto. El mar retropalpebral tiene una parte tras el párpado superior y otra bajo el inferior. A su vez, el mar lacrimal expuesto tiene una cisterna, un lago, unos ríos y una película prebulbar. El mar retropalpebral (mare retropalpebrale) está limitado por dos superficies lisas (la palpebral y la bulbar). En la parte ocupada por el fondo de los fórnices se amolda a los pliegues conjuntivales de la zona, y en la parte prebulbar es una fina lámina capilar, que alcanza su mínimo espesor cerca del margen palpebral superior. La estructura grosera del mar retropalpebral consta de 3 capas : la anterior y posterior mucosas y la intermedia acuoserosa. El mar lacrimal retropalpebral superior tiene una lágrima más seromucosas y de características nacientes que el inferior. El mar retropalpebral inferior tiene más mucina y en él se encuentran grumos y filamentos mucosos, probablemnete por sedimentación de la mucina del resto de mar, donde se degrada y se elimina. La cisterna lacrimal (cisterna lacrimalis) es la porción piramidal triangular del menisco lacrimal alojado en el triedro formado entre el bulbo ocular y el ángulo externo de los dos párpados (Murube 1981a, p 751). Los ríos lacrimales (rivi lacrimales) son los meniscos de lágrima formados a lo largo de los párpados superior e inferior. El lago lacrimal (lacus lacrimalis) es la parte triangular del mar lacrimal expuesto comprendida entre los puntos lacrimales y el canto interno. El contenido del lago lacrimal está aumentado por la presencia de los pliegues lacunares. La película lacrimal interpalpebral o PLIP (pellicula lacrimalis interpalpebralis prebulbaris) es la lámina de lágrima situada delante de la superficie ocular expuesta, entre los meniscos de la cisterna, los ríos y el lago lacrimal. Cuando el ojo está abierto, la superficie de la película lacrimal es de 1'5-2 cm2. El espesor de la película precorneal se calculó primero por la colocación de una fibra de vidrio dentro de la película (Mishima 1965), o la colocación de un pequeño disco absorbente sobre un área de la película y su posterior pesado (Ehlers 1965) o por diversos métodos de tinción/dilución, encontrándosele un espesor de entre 3 y 10 m m : 3-4 m m (Norn 1966), 7 m m (Holly et al 1971b) o 6-10 m m (Ehlers 1965b). Pero recientemente, Prydal 1992ab, usando métodos de láser-interferometría y de microscopía de escaneo confocal, ha encontrado valores de 40 m m, de los que los 30 m m más profundos son mucus y los 10 m m más superficiales son acuoserosos . En la rata, Anderton 1996 encuentra que el espesor es de 36 m m (rango 23-70 m m), y que si instila en la cuenca lacrimal un mucolítico, baja a 19 m m (rango 10-34 m m). Cualquiera sea el espesor, parece que es máximo al 1-2 segundos de abir los párpados, y a partir de ese momento disminuye por evaporación. Tiene un estrechamiento longitudinal vecino y paralelo a cada menisco lacrimal que, como se ve obscuro cuando se mira por biomicroscopia ultravioleta despues de haber instilado una gota de fluoresceína, se llama línea negra (McDonald et al 1971) Que la película lacrimal no es una fase homogénea sino que tiene tres grupos de componentes distintos -acuoseroso, mucoso y lipídico- ya fué escrito por Terson 1892 ("du mucus est fourni par les sillons à epithélium caliciforme, quelques produits surtout sébacés par la caroncule, une sécrétion séreuse par les glandes lacrymales"), y Schirmer hizo alusión a una cierta estratificación al señalar que al absorber el líquido del mar lacrimal queda sobre la conjuntiva una fina capa de moco ("ein zartester Schleimbezug"). Pero la primera persona que claramente señaló que hay un estrato lípido anterior, uno acuoseroso medio, y uno mucoso posterior, fué Wolff (1946, 1954), y su concepto ha servido de hipótesis de trabajo durante medio siglo. Sin embargo, el concepto de la triestratificación de la película lacrimal está actualmente en revisión: La película lacrimal tal vez no sea un triestrato de lípidos, agua y mucina, sino un gel de mucina con una capa lípida flotando sobre él. Cuando la película lacrimal se adelgaza por evaporación, la capa lípida se acerca a la mucina e interacciona con ella. Los lípidos meibomianos se combinan especialmente con los fosfolípidos mucínicos, formando compuestos hidrófobos, a cuyo nivel, si un nuevo parpadeo no lo impide, aparece un islote de desecación que rápidamente se extenderá. En la interacción lípidos-mucina participa, facilitándolo, el flujo convectivo Marangoni-Bénard (Velarde et al 1986). La capa lípida de la película lacrimal, cuando el ojo está abierto, ocupa en superficie todo el área interpalpebral expuesta, y en espesor mide 0'05-0'10 m m. El espesor de la capa lípida, según Korb et al 1996, es de 0'079 m m en el humano y 0'220 m m en el conejo. La medida del espesor de la capa lípida por interferometría cromática se ha hecho muchas veces (ver capítulo 25 de este libro). Cuando lo párpados se cierran , las moléculas lípidas se montan unas sobre otras y la capa puede alcanzar 1 m m o más. Cuando los párpados se abren, lo hacen a una velocidad de 8 cm/sec. (Berger et al 1974), y como la capa lipída se extendería más rápidamente -aproximadamente a 25-27 cm/sec-, nunca queda lágrima acuoserosa descubierta La capa lípida se forma mayoritariamente por la secreción de las glándulas de Meibomio, y es posible que participen en pequeña cuantía las glándulas de Zeis y de Moll. La capa lípida de la película lacrimal contiene un 80-90% de lipidos de baja polaridad, que incluyen principalmente ésteres de cera y de colesterol y trazas de triglicéridos; estos componentes de baja polaridad se sitúan en la parte anterior de la capa. El restante 10-20% son lípidos de alta polaridad, que incluyen ácidos grasos libres, alcoholes alifáticos, glicolípidos y pequeñas cantidades de lecitinas y otros fosfolípidos estos componentes de alta polaridad se sitúan en la parte profunda de la capa, orientándose sus moléculas paralelas unas a otras, con su grupo polar hacia la fase acuoserosa y la larga cadena hidrocarburada hacia los componentes lípidos de baja polaridad. Las funciones principales de la capa lípida son : (1) Dificultar la evaporación de la lágrima acuoserosa, cosa ya expuesta por Wolff 1946. (2) Presionar a la capa seromucosa y ayudarla a que se extienda sobre la capa mucínica. (3) Por esa misma presión, que aumenta al engosar su espesor al cerrar los párpados, evitar que la lágrima acuoserosa se derrame en forma de una gota al parpadear. Cuando el ojo está abierto, si la capa lípida tiene un espesor de 0'16 m m ejerce una presión de película sobre la fase acuoserosa de 10-12 dinas/cm. La capa acuoserosa de la película lacrimal está constituida fundamentalmente por la secreción de las glándulas lacrimales principal y accesorias, y tal vez en una pequeña proporción por líquido atraído por la hiperosmolaridad de la lágrima de las paredes de la cuenca lacrimal. La capa mucosa de la película lacrimal procede mayoritariamente de las células mucíparas, y minoritariamente de la mucina segregada por las glándulas lacrimales. Se adsorbe esta mucina al glycocalyx o mucina epitelial, que es una arborización glicoproteica que las células epiteliales no secretorias de la córnea y la conjuntiva desarrollan hacia el exterior como parte de su membrana celular superficial. Una de las funciones de la mucina es cubrir el epitelio corneal, que es dacriofóbico, con una capa glicoproteica que lo haga dacriofílico y permita a la lágrima extenderse sobre él. Un líquido se extiende sobre un sólido cuando la suma de la tensión superficial del líquido más la tensión interfásica líquido-sólido es igual o menor a la tensión superficial crítica del sólido. La tensión superficial crítica del sólido, -en nuestro caso, el epitelio corneal- es de unas 28 dinas/cm. Por lo tanto la superficie corneal no puede ser mojada por el agua, cuya tensión superficial a 35ºC es 70 dinas/cm. Para salvar esta diferencia, por un lado el parpadeo unta de mucina la superficie corneal y eleva su tensión superficial crítica a aproximadamente 38 dinas/cm; y por otro lado se disuelve en la fase acuoserosa mucus y rebaja su tensión superficial a aproximadamente 38 dinas/cm. Así, la fase acuoserosa se puede extender sobre la superficie corneal, aunque en un equilibrio inestable. Este equilibrio adquiere mayor estabilidad al estar la capa acuoserosa bajo la presión de membrana de la capa lípida.
PARPADEO Los movimientos palpebrales fundamentales son 3 : El parpadeo, la connivencia y el blefarospasmo. El parpadeo fisiológico para ser tal debe ser inconsciente. El parpadeo voluntario consciente se acerca más a un blefarospasmo sui generis que a un parpadeo fisiológico. El parpadeo fisiológico tiene dos formas que se suceden : parpadeos incompletos y completos. En los primeros, los párpados no llegan a tocarse, no se da fenómeno de Bell y se producen por secuencias o trenes de 4-9 parpadeos seguidos. En los segundos, los párpados llegan a contactar, se esboza el fenómenos de Bell y se dan por secuencias de 1-2 parpadeos seguidos (Murube 1981a p 451). Cada parpadeo incompleto dura por término medio 0'225 sec y el completo 0'250 sec (p 452). La frecuencia del parpadeo es de 5-10 veces/minuto (Schirmer 1903). La más reciente medida de la frecuencia del párpadeo en humanos normales es de 8'58 parpadeos/minuto, con un intervalo de 7'01 sec entre cada parpadeo (Korb et al, 1996). El desplazamiento de los párpados (figura 2-13) durante el párpadeo se hace en las tres dimensiones : verticalmente, medialmente y hacia atrás. (1) En cuanto al desplazamiento vertical, al cerrar los ojos, el espacio interpalpebral se cubre a expensas del párpado superior que desciende 9-9'5 en el centro y 2-3en el canto lateral el párpado inferior no sólo no asciende, sino que desciende 2-3 mm en el cantus lateral y 0'5 mm en su punto tangente a la córnea. (2) En ambos párpados, al desplazamiento vertical se añade un desplazamiento medial, que es máximo, de unos 2 mm, a nivel de los puntos lacrimales. (3) El desplazamiento posterior es de 1mm en el el área que queda sobre el punto medio de la córnea para el párpado superior, y practicamente nulo en igual sitio del párpado inferior. La presión que ejerce el párpado superior al descender sobre el globo se hace principalmente con el borde inferior del tarso. Su presión se evalúa en 10-15 g en una persona joven y hace hundirse al globo ocular en la órbita. Esta presión parece ser muy importante para la liberación de los lípidos meibomiano, porque como determinaron Chew et al 1993, en ausencia de parpadeo, estos lípidos no se vierten al borde palpebral. Según nuestras observaciones este desplazamiento juega también un papel importante en el flujo lacrimal, pues tensando los ligamentos cantales y desplazando el globo hacia atrás, crea una zona de menor presión en los cantos, especialmente en el lateral, de forma que tras cada párpadeo se ve que ha fluido una importante cantidad de lágrima retropalpebral a la cisterna lacrimal.
El fenómeno de Bell, según fue descrito por este autor en 1823, es la elevación del ojo a cada parpadeo normal. La razón del fenómeno de Bell es deconocida. Mackie (1971) propuso que sería el facilitar el untado de mucina rotando el ojo hacia arriba. Nosotros (Murube 1981a, p 434) propusimos que, como esta sursumducción se hace a expensas de los rectos inferiores (no sería lógico pensar que la hacen los rectos superiores precisamente cuando el músculo elevador del párpado superior se relaja) sería empujar el ojo hacia adelante y dificultar su retroceso para que la presión ejercida por el párpado sea más eficaz. El cierre palpebral de connivencia (del latín connivere, cerrar los ojos) es la oclusión palpebral mantenida para evitar la estimulación lumínica de la retina, y para proteger mantenidamente la córnea. Es más lento que el movimiento de parpadeo, presiona menos el globo ocular y apenas esboza en su primer momento un desplazamiento medial de los párpados, por lo que no bombea lágrima. La connivencia ocular es frecuente en pacientes de ojo seco para disminuir las molestias de desecación. El blefarospasmo adopta diversas formas : espasmos mantenidos, parpadeos espásticos, tics, temblores, trémores y parpadeos voluntarios. En pacientes de ojo seco son frecuentes los parpadeos espásticos y los parpadeos voluntarios.
FLUJO DEL MAR LACRIMAL El mar lacrimal tiene un flujo que se establece durante y tras el parpadeo. Durante el parpadeo, la lágrima acuoserosa, producida casi todas ella en el fórnix superior, es arrastrada y exprimida hacia abajo, especialmente hacia la cisterna lacrimal. La lágrima situada tras el párpado inferior es empujada hacia el lago lacrimal y hacia el rivus inferior. La lágrima de la película lacrimal y los meniscos lacrimales se funden parcialmente en el espacio prismático interpalpebral del cierre. La mucina del espacio retropalpebral superior es extendida y depositada sobre la córnea y la conjuntiva expuesta, y la mucina degrada existente en la superficie expuesta es arrastrada hacia los meniscos y el fórnix inferior. La capa lípida va montando sus capas moleculares una sobre otra, y empujada hacia adelante por la capa acuoserosa que también engruesa al cerrar los ojos, llega a contactar con las glándulas lípidas anteriores, contaminándose con ellas. Al abrir los párpados, el párpado superior arrastra hacia arriba su propio menisco lacrimal acuoseroso y deja otro menisco en el párpado inferior. La película lacrimal se va reformando, manteniendo durante unos segundos una corriente ascendente de la fase acuoserosa por la succión del menisco superior que es inicialmente deficitario, y otra corriente ascendente de la capa lípida, que busca una distribución uniforme. Tras el parpadeo, la lágrima situada bajo el párpado superior, impulsada por la presión palpebral, la gravedad, la vis a tergo de nueva lágrima y eventualmente la succión de los meniscos lacrimales deficitarios, desciende hacia el menisco superior lateral y por él baja lateralmente hasta la cisterna, y otra fracción pasa directamente a la cisterna desde el espacio retropalpebral. Nosotros (Murube 1981a, p 643) hemos precisado la existencia de un pliegue mucoso horizontal casi constante situado en la conjuntiva palpebral bajo el ligamento cantal lateral, por donde la corriente retropalpebral aflora con fuerza en la cisterna lacrimal esto es fácilmente comprobable tiñendo con fluoresceína la cisterna y viendo como aflora en ella un chorro de lágrima no teñida proveniente de detrás del párpado, o viceversa, poniendo un papel teñido de fluoresceína bajo la parte lateral del párpado superior y viendo como entre parpadeo y parpadeo emerge un chorro amarillo subacuático que fluye desde el pliegue descrito a la cisterna lacrimal. La cisterna se nutre principalmente de lágrima que le llega del espacio subpalpebral vecino y del río superior lateral. La curvatura de la concavidad de la pared anterior del menisco es una constante dependiente de fuerzas de superficie y volumen. Cuando por exceso de lágrima esta superficie se hace menos cóncava o se aplana, tiende a recuperar su curvatura ideal, eliminando el exceso lacrimal hacia los ríos. Por el contrario, cuando por falta de lágrima la superficie anteroexterna del menisco se hace excesivamente cóncava, tiende a recuperar su curvatura tomando lágrima de las inmediaciones, especialmente del espacio subpalpebral. Así, la cisterna lacrimal es no sólo un depósito, sino un regulador de la lágrima del mar lacrimal. La película lacrimal queda fuera de fuerzas dependientes de los párpados, que están quietos, o de la gravitación, pues como expresó Ehlers 1965b, por ser tan fino su espesor, sus fuerzas de superficie son tan superiores a las gravitatorias, que el espesor de la película es aproximadamente el mismo en su mitad superior y en su mitad inferior. Por el contrario, desde que se abren los ojos la película lacrimal sufre un cambio precoz, rápido y continuo, dependiente de la evaporación del agua y de la interación entre las capas lípida y acuoserosa. El río inferior, que aumento al cerrar el ojo con la lágrima de la película palpebral y del río superior, y se adelgazó al abrir los párpados cediendo lágrima a la película lacrimal y al lago lacrimal, se nutre al dejar el ojo abierto principalmente de la cisterna lacrimal y fluye hacia los puntos lacrimales, empujado por la vis a tergo procedente de la cisterna y trasmitida por el menisco encerrado bajo la capa lípida del menisco, y por la succión del canalículo tras el arpadeo (fenómeno de Krehbiel, 1878). La cisterna lacrimal recibe lágrima acuoserosa de todas las áreas vecinas, especialmente del río inferior y del espacio retropalpebral inferior. Recibe también mucina dispersa en la lágrima o conglomerada en grumos que engloban células epiteliales, leucocitos, hematíes, gérmenes y cuerpos extraños. Recibe así mismo lípidos de la capa lípida de la película lacrimal y de las glándulas lípidas carunculares; en el lago lacrimal hay a menudo burbujas de envuelta lípida y contenido desconocido, y gotitas grasas. La existencia de glándulas de secreción lipídica en la carúncula, no tiene hasta el presente explicación probada. Nosotros propusimos (Murube 1981, pp 431,466) que una de sus funciones prodría ser degradar el mucus lacrimal por contaminación lípida, interaccionando con la capa oleosa de la película por delante, y con la secreción de las glándulas lípidas carunculares, por detrás esto y la acción enzimática de la lágrima, fragmentaría los complejos mucolípidos y, haciéndoles perder su tendencia adsorbente al epitelio, facilitaría su expulsión por los puntos lacrimales.
ELIMINACIÓN DEL MAR LACRIMAL El recambio o índice de renovación (turnover) normal del mar lacrimal, por fluorofotometría, es del 16% / minuto (Mishima et al 1966), del 10% (González de la Rosa et al 1981), o del 17'32% / minuto (Benítez del Castillo et al 1993). Por un método con isótopos radiactivos, Sörensen et al 1979 lo evaluaron en 8'3 % / minuto. La desaparición del mar lacrimal se hace por evaporación, por absorción por las paredes de la cuenca lacrimal y por emunción por la vía lacrimal. Evaporación. Ya Bernard 1843-5 y Santos Fernández 1903 señalaron que los pacientes con oclusión de vías lacrimales sin epífora eliminan su mar lacrimal por evaporación. Hyrtl 1847 creía que en condiciones normales toda la lágrima basal se elimina por evaporación, y que las vías lacrimales sólo funcionan en caso de hipersecreción. Y Charlín Correa 1925 consideraba que la evaporación era la forma cuantitativamente más importante de eliminación lacrimal. El primer cálculo cuantitativo de la evaporación lacrimal se debe a Schirmer 1903, que la evaluó calculó en 0'27g de agua al día. Posteriormente se ha calculado en 2'2-3'7 m l/cm2/hora (o Mishima et al 1961, o Maurice 1961, o von Bahr 1941), 3'633 mg/cm2/hora (Iwata et al 1969), 8'7 mg/cm2/hora (Hamano et al 1980). Si se toman como valores normales de producción lacrimal el de 1'2 m l/min y como de evaporación en atmósfera con 40 % de humedad el de 0'1 m l/min, resulta que el 8% del volumen del mar lacrimal desaparece por evaporación. La evaporación de la película lacrimal de un paciente de ojo seco es 2 veces más rápida que la de un ojo normal (Rolando et al, 1983). Si falta totalmente la capa lípida, la evaporación aumenta 4 veces (Iwata et al 1969) o 20 veces (Mishima et al 1961). Absorción parietal. La cantidad de lágrima eliminada a través de las paredes de la cuenca lacrimal ha sido calculada en ojos humanos normales en 0'024/min (Meyer et al 1975), 0'0176/min (Denffer et al 1979) y 0'0145/min (Sörensen et al 1979b) Emunción por la vía lacrimal. En condiciones normales, parte del mar lacrimal se elimina a través de los puntos lacrimales hacia el saco lacrimal, donde parcialmente se absorbe y parcialmente se elimina hacia el conducto lacrimonasal . En condiciones de sobrecarga, la vía lacrimal es el principal camino de eliminación.
FUNCIONES DEL MAR LACRIMAL Óptica. Formar una película precorneal muy lisa, gracias a la baja tensión superficial del líquido lacrimal. Esto es importante, pues la superficie anterior de la córnea es el principal dioptrio del ojo. Cuando el ojo está seco y el epitelio queda en contacto con el aire, su irregularidad hace que las imágenes se vean desenfocadas y borrosas. Metabólica. Aportar al tejido ectocorneal agua, oxígeno, glucosa, reguladores,enzimas, etc. Por ej., la b -amilasa lacrimal, actúa en la endohidrolisis del glucógeno de las células del epitelio corneal, liberando así la glucosa necesaria para su metabolismo. Limpiadora. La lágrima es el vehículo que elimina detritos epiteliales, cuerpos extraños y microorganismos. Si la lágrima faltase, estas materias quedarían retenidas en la cuenca lacrimal. Antimicrobiana. Por: (1) barrera y barrido mecánico, (2) enzimas de la lágrima con un cierto efecto bacteriostático y bactericida : lactoferrina, aril-sulfatasa A, peroxidada, lisozima, (3) fracciones inmunológicas específicas ligadas a las Ig lacrimales y (4) células de las defensas inespecíficas y específicas que pasan a la lágrima. Lubricante. La lágrima, debido a su adsorción a la superficie de la cuenca lacrimal y a su escasa viscosidad, es un buen lubricante para el deslizamiento de párpado superior durante el parpadeo. Otras funciones secundarias, como la fotoabsorbente, la reflexiva de la luz, la aclarante, la tamponadora y la humectante nasal, son accidentales.
COMPONENTES DEL MAR LACRIMAL Se considera que de la lágrima existente en el mar lacrimal, el componente acuoseroso es aproximadamente el 99'78 % de la lágrima y se produce en las glándulas lacrimales y en pequeña parte proviene del corion mucoso y de la córnea traspasando el epitelio de la cuenca lacrimal; el 0'2 % es mucina que procede de las células caliciformes; y el 0'02 % es grasa que procede de las glándulas lípidas. |