HISTORIA Y HUMANIDADES
La osmolaridad a ambos lados de la
córnea
MURUBE J, RIVAS L, CAMBLOR JP, CORREA C,
MUÑOZ-NEGRETE F, SALES-SANZ M, CABREJAS L
El presente trabajo ha sido desarrollado
como parte del programa 2005-2008 de la Fundación de Investigación Médica Mutua
Madrileña.
El conocimiento de la salinidad de los líquidos y su influencia
en los procesos físicos y biológicos ya se inició de antiguo. Ya en el siglo I,
el romano-cordobés Séneca (1) escribió que las aguas saladas son más
pesadas, y que descienden, mientras las menos saladas ascienden. Pero la primera
referencia que a nuestro saber se hizo sobre los líquidos oculares salinos se
refieren a la lágrima, y así, el greco-romano Galeno (2) (siglo II, AD)
dijo que la xeroftalmía es una enfermedad ocular con lágrima salobre y nitrosa,
y unos siglos más tarde, el griego Aecio de Amida (3) (siglo V AD)
escribió que la psoroftalmía es un enfermedad de la superficie ocular con
lágrimas salinas, corrosivas, y úlceras cantales.
Las bases de los actuales conocimientos sobre la osmolaridad de
los tejidos vivos se iniciaron en 1748, cuando el abad francés Jean-Antoine
Nollete (4), que a la sazón trabajaba como profesor de Física Experimental
en la Universidad de Navarra, observó que en un recipiente con agua azucarada a
un lado de una membrana orgánica y agua pura o insípida al otro lado, el agua
pasaba del lado insípido al azucarado, pero no a la inversa, de forma que el
agua azucarada aumentaba de volumen y se hacía menos concentrada, mientras que
el agua pura disminuía de volumen y se mantenía insípida. Este experimento
inicial se repitió con diversas membranas y líquidos.
Dutrochet (5) fue el primero que en 1828 inició el concepto
de membrana semipermeable, es decir, aquélla cuyos finos poros permiten el paso
a su través del agua, pero no de los iones de sal, y por ello es permeable al
solvente acuoso, pero no a los solutos. La perfusión ocurre siempre desde la
solución acuosa menos concentrada a la más concentrada. Llamó a esto «ósmosis»,
neologismo derivado del griego wsmóç, acción de
empujar, y «presión osmótica», a la fuerza con la que el líquido menos salino
impulsaba su agua al más salino. El flujo osmótico cesaba cuando ambos líquidos
alcanzaban la misma concentración iónica a ambos lados de la membrana, o cuando
el más concentrado alcanzaba una mayor altura de nivel, cuyo peso frenaba el
flujo. Así, la presión osmótica se pudo medir por la diferencia de nivel final
de los líquidos a ambos lados de la membrana.
A mediados del siglo pasado, el botánico alemán van Mohl
(6) observó que las membranas de las células vegetales y animales eran
semipermeables, y tratando de explicar el cambio de fluidos a su través, dio la
primera explicación científica de la ósmosis. El también botánico alemán
Pfeffer (7) (1877) estudió la presión osmótica poniendo agua azucarada con
sacarosa en un recipiente de arcilla, que a su vez colocó en un recipiente mayor
lleno de agua pura, y midiendo los cambios finales de altura de los niveles de
agua, determinó por primera vez con un manómetro la presión osmótica.
La medida de la presión osmótica por el método de Pfeffer tarda
bastante porque el flujo osmótico es lento. Hartley (8), trató de
acelerar las medidas osmométricas aplicando al líquido hipertónico una presión
que detuviese el flujo a través de la membrana semipermeable. Pero el mayor
adelanto se hizo cuando Raoult (9) publicó en 1882 que la congelación de
un líquido solvente es tanto más baja cuanto más electrolitos tiene, pues el
punto de congelación depende de la osmolaridad y no de la naturaleza de los
solutos. El punto de congelación del agua pura es por definición 0°C, y el del
suero humano es 0,532 a 0,539°C, lo que significa que tiene una osmolaridad de
285 a 290 mOsm/l. Otros métodos osmométricos son la determinación del punto de
rocío o de temperature de condensación del vapor, la presión de vapor, la
conductividad eléctrica, etc. pero en los laboratorios de investigación clínica,
la determinación del punto de congelación sigue siendo el método más usado.
Se conocían de antaño los niveles de salinidad de mares y
fuentes, y esto se relacionó enseguida con la ósmosis y su repercusión en
plantas y animales. Los océanos abiertos tienen una salinidad que ronda los 35 g
de NaCl (y en menor proporción , de otras sales) por litro, es decir, 4 veces
más que la media de los fluidos orgánicos de los seres humanos. Hay mares que
reciben aguas fluviales generalmente poco salinas, y en los que la evaporación
es escasa, y por tanto tienen una baja salinidad, como algunas áreas del mar
Báltico, que alcanzan una salinidad de sólo 5 g NaCl/l, casi la mitad de la de
la sangre humana. Hay otros mares que reciben aguas fluviales pero que no
desaguan a los océanos, sino que lo hacen por evaporación, y por tanto tienen
una alta salinidad, como el mar Muerto, que alcanza 340 NaCl/l, casi 40 veces
superior a la del suero humano. En las aguas de fuentes y manantiales, la
salinidad no depende de la evaporación , sino de la disolución de las sales
subterráneas por las que pasan, y así, el agua de las salinas de Leniz, en
Guipúzcoa, tiene una salinidad que le da una osmolaridad equivalente a 85g/l de
NaCl.
Estos conocimientos desarrollaron un gran progreso en la
comprensión de la biología vegetal y animal, y en muchas actitudes médicas, una
de las cuales reseñamos como curiosidad, pues su conocimiento de hace más de un
siglo sigue teniendo aplicación práctica hoy día: el sujeto casi ahogado por
inmersión, tiene sus alvéolos pulmonares llenos de agua y perecerá por asfixia
si no se trata de inmediato: si la inmersión fue en agua de piscina, debe ser
tratado directa y urgentemente con respiración artificial boca a boca, pues el
agua que llena sus alvéolos pasa por ósmosis en sólo minutos a su cuerpo; si la
inmersión fue en agua marina, el agua intraalveolar no sólo no se marcha, sino
que atrae más agua del cuerpo del paciente, por lo que cualquier respiración
artificial debe ir precedida del urgente vaciamiento del agua pulmonar por
compresión torácica con la cabeza baja, y ladeada o prona.
En los fluidos orgánicos animales la tonicidad habitual
ronda en el suero sanguíneo una equivalencia a una solución de NaCl de 9g/l, lo
que le da una osmolaridad de 285-290 mOsm/l. La osmolaridad del líquido
cefalo-raquídeo es de 290-295 mOsm/l, la de la saliva total 200-310 mOsm/l, y la
de la orina, 200-500 mOsm/l. Los líquidos que bañan las caras posterior y
anterios de la córnea, es decir, el humor acuoso y la lágrima, tienen una
osmolaridad de 300 a 310 mOsm/l.
PLASMA SANGUÍNEO, HUMOR ACUOSO Y LÁGRIMA
El humor acuoso tiene un volumen de aproximadamente 0,06 ml en
cámara posterior y 0,25 ml en cámara anterior. Su flujo es de aproximadamente 2
µl/minuto. Contiene sales, siendo la principal el NaCl, pero teniendo también K,
Ca, fosfatos y bicarbonatos. Asimismo tiene componentes no electrolíticos o
escasamente electrolíticos, de los que unos son más escasos que en sangre, como
la glucosa (0,6%° en acuoso y 1%° en sangre) y otros más abundantes, como el
ácido ascórbico (15%° en acuoso y 0,05%° en sangre). Como resultado de todo
ello, el humor acuoso es hiperosmolar con respecto al suero sanguíneo. Esto
juega un papel en su formación y eliminación (10).
El mecanismo de formación del humor acuoso tiene dos componentes
principales: la ultrafiltración y la secreción. En el mecanismo de
ultrafiltración juega un papel importante la presión hidrostática sanguínea
en los capilares ciliares (² 60 mmHg) que permite que su contenido pueda ser
empujado a través de sus capilares fenestrados, estroma y epitelio ciliar hacia
la cámara acuosa posterior, aunque condicionado por otros factores, entre otros,
que las substancias permeantes no son uniformes, ya que por ejemplo la urea es
menor en acuoso que en plasma (11).
En el mecanismo secretor juega la osmolaridad un
importante papel. Los espacios intercelulares del epitelio ciliar funcionan como
canales intercelulares, y el líquido vertido a ellos por las células no
pigmentadas del epitelio ciliar fluye hacia la cámara posterior. Estas células
claras, merced a la energía para el trasporte iónico activo suministrada por su
ATPasa, vierten Na+ a los canales intercelulares del epitelio ciliar
y a la cámara posterior, subiendo su osmolaridad. Es el fenómeno clave de la
secreción de acuoso (12). Este acuoso incipiente alcanza en la parte más
profunda de los canales intercelulares una osmolaridad de 335-340 mOsm/l, la
cual atrae agua, y su osmolaridad va reduciéndose hacia el extremo de los
canales más cercanos a la cámara posterior.
El humor acuso tiene siempre una osmolaridad superior a la del
suero (13,14), sufriendo numerosas variaciones circunstanciales, y así
Schaeffer (13) encontró en humanos que el acuoso es algo hiperosmolar en
relación con el plasma, pero que en las anestesias generales la osmolaridad
sanguínea sube algo y se iguala a la del acuoso
Este humor acuoso, con una osmoralidad media de 305 mOsm/l, se
elimina principalmente a través del conducto de Schlemm y de la úvea, pero
también en escasa cuantía puede hacerlo a través de la córnea. con determinados
componentes selectivos por factores de pinocitosis y de presión hidrostática. En
un estudio experimental Ridley (15) observó que el humor acuoso, si se le
aplica una presión de 25 mmHg, puede atravesar la córnea a un ritmo de 0,81
µl/min, y si de 250 mmHg, el flujo sube a 1,13 µl/min. En el individuo normal,
la subida de presiones intraoculares sobre 50 mmHg provoca un edema corneal
clínicamente detectable.
Cogan et al (16), en 1942, emitieron la teoría de que el flujo
fisiológico de agua en la córnea entra por su estroma limbal. El estroma corneal
es fuertemente hidrófílo, debido a sus glucosa-amino-glicanos, especialmente a
su componente keratán sulfato. La deturgescencia corneal se hace hacia el acuoso
a través del endotelio corneal, y hacia el mar lacrimal a través del epitelio
corneal. La figura original de Cogan esquematiza este flujo (fig. 1).

Fig. 1. Esquema del flujo de fluido a través de la córnea. Las flechas indican
la dirección de la corriente, y la densidad de las flechas desde la periferia
hacia el centro indica el movimiento relativo de la cantidad de flujo. La
cantidad de flujo hacia las superficies anterior y posterior no está aún bien
conocida (Cogan et al 1942) (16).
La cuantificación del flujo intracorneal fue motivo de numerosos
estudios durante los años siguientes,y ello ha complementado y modificado en
muchos aspectos el clásico esquema de Cogan y Kinsey. Brubaker et al (14)
(1962) calcularon en conejos la tonicidad del suero sanguíneo, humor acuoso, y
líquido intersticial de la córnea periférica o limbal, y de la córnea central o
prepupilar, y encontraron valores de osmolalidad respectivamente de 294,3-294,9
mOsm/kg, 305,1-306 mOsm/kg, 302,8 y 305,3 mOsm/kg. Aparte de la osmolaridad y de
la presión intraocular, otros factores importantes son la integridad anatómica
del endotelio y epitelio y el metabolismo corneal (17).
Las células del endotelio y epitelio corneales son ricas en
lípidos, por lo que son más fácilmente atravesables por los componentes
liposolubles, mientras que el estroma corneal es más fácilmente permeable a los
acuosolubles. Las barreras ricas en lípidos son fácilmente atravesables por los
compuestos no polares, mientras las ricas en agua, lo son por los compuestos
polares.
El endotelio corneal juega una función de barrera y otra de
bombeo, pasando diversos componentes en ambas direcciones, pero principalmente
agua, sodio y otros electrolitos desde el estroma corneal hacia cámara anterior,
y dando como resultado final una acción deturgescente de la córnea.
El epitelio corneal tiene unos factores comunes y otros
diferenciales con el endotelio. La hiperosmolaridad relativa de la lágrima hace
que continuamente pase agua de la córnea y de la conjuntiva bulbar a la película
lacrimal (15,16,18-25), Michel (26) (1884) fue el primero en exponer que
la evaporación de la película lacrimal juega un importante papel en aumentar su
osmolaridad y en atraer agua de los tejidos limítrofes. Y 3 años despuués,
Massart (27) (1887) determinó su influencia en la deturgescencia de la
cornea y la disminución de su espesor. Mishima (28) (1965), observó que
los conejos sólo parpadean unas pocas veces durante una hora, por lo que la
evaporación de su película lacrimal es alta, y que sin embargo su tonicidad se
mantiene equivalente a una solución acuosa de NaCl al 9%°, y supuso este autor
que en ellos hay una transferencia transcorneal de humor acuoso que evaluó en 6
µl/hora.
La tonicidad de la lágrima se ha medido desde hace siglo y medio
con métodos cada vez más exactos, dándole los valores que se expresan a
continuación (tabla 1).

En los síndromes de ojo seco, la lágrima suele ser hiperosmolar
respecto a la del individuo sano, lo que fue primero detectado por Mastman et
al (42) (1961), y posteriormente confirmado por muchos autores. La tonicidad
media de la lágrima en los síndromes de ojo seco, según diversos investigadores,
es la siguiente (tabla 2).

La osmolaridad del mar lacrimal puede modificar en sentido
positivo o negativo el flujo acuoso a través del epitelio corneal según sus
propias características bioquímicas, y las característica de este epitelio.
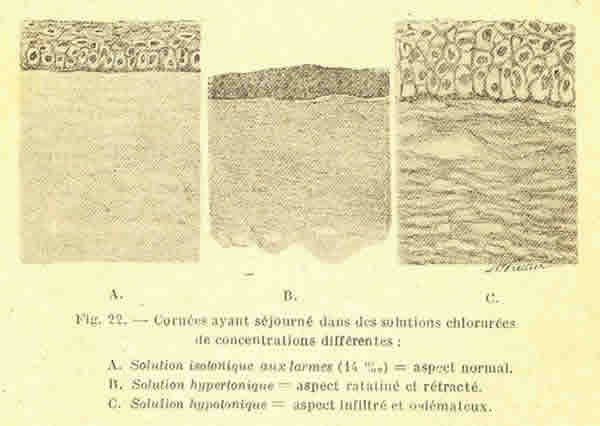
Reproducimos la primera imagen que recoge este hecho, publicada por Cantonnet
1925, pues creemos que pese a su interés nunca ha sido reproducida en la
literatura posterior hasta el presente trabajo (fig. 2).


Fig. 2. Córneas que han permanecido en una solución clorurado-sódica de
concentraciones diferentes.
A. Solución isotónica a la lágrima (14%). * = aspecto normal.
B. Solución hipertónica = aspecto reseco y retraído.
C. Solución hipotónica = aspecto infiltrado y edematoso.
* Según la valoración de Cantonnet.
Los colirios hiperosmolares pueden tener una acción sobre el
flujo de humor acuoso. Sin embargo no se han usado para disminuir la presión
intraocular por su escasa duración y eficacia. Así, el tiempo de duración en el
mar lacrimal de un colirio hiperosmolar es muy escaso. Mastman et al (42)
(1961) determinó que el mar lacrimal al que se instila un colirio con un 25%°de
NaCl recupera la normoosmolaridad en sólo 1 1/2 a 2 minutos, siendo ello
principalmente debido no a la atracción del agua intracorneal, sino a la
hipersecreción lagrimal refleja que estimula. En el mismo sentido, Sust Mir
(63) (1967) ratificó que la persistencia de un colirio hiperosmolar es muy
pasajera. Y Hollyet al (64) (1981) instilaron colirios hipertónicos de
NaCl de 500 y 1.000 mOsm/kg, y observaron que en pocos minutos se recuperaba la
normotonicidad del mar lacrimal.
Los colirios hiper e hiposmolares alteran el epitelio de la
superficie ocular. Concomitantemente tienen una acción deturgescente corneal
según ya mostraron Massart (27) (1889) y Cantonet (32,33,65)
(1908, 1925), y los hipoosmolares, una acción edematizante. Concomitante a ello,
alteran el epitelio de la superficie ocular (fig. 3). Von Bahr (22)
(1956) determinó que una solución de NaCl del 14%° produce en 10 minutos un
adelgazamiento corneal del 15,9%. Este efecto deshidratante se acompaña de un
menor o mayor daño epitelial (45,66-68). Entre las acciones deletéreas sobre el
epitelio de la superficie ocular están la destrucción de células caliciformes de
la conjuntiva, en las que los trabajos de Egbert et al (67) y Vergés
(68-71) fueron pioneros. Por ello los colirios y pomada hiperosmolares no se
usan para achicar humor acuoso y reducir la presión intraocular, sino sólo para
reducir momentáneamente los edemas corneales, usándose generalmente pomadas o
colirios con concentraciones de NaCl al 50%°.

Fig. 1. Citología de impresión de córneas de rata,
después de instilarles cada 5 minutos un colirio, durante media hora. Tinción
PAS-hematoxilina. Aumento original x40.
A. Colirio isotónico a la lágrima (10%). Las células epiteliales tienen los
bordes bien marcados. El citoplasma es intensamente eosinófilo. Los núcleos
están bien manifiestos.
B. Colirio hipertónico. Las células epiteliales tienen los bordes celulares muy
definidos. El citoplasma está muy poco teñido, siendo ligeramente eosinófilo.
Los núcleos aparecen poco marcados.
C. Colirio hipotónico (agua destilada). Las células epiteliales tienen los
bordes celulares muy bien marcados. El citoplasma es poco eosinófilo
(ligeramente basófilo). Los núcleos están moderadamente manifiestos.
BIBLIOGRAFÍA
-
Séneca LA. Naturales Quaestiones. Liber I, Capt 6. Circa 62
AD.
-
Galeno. E Iatroz. Liber XIV. Siglo II AD.
-
Aecio de Amida (Siglo V) Tetrabiblion, cap. II (Enfermedades
oculares), LXXVII (escleroftalmía), LXXVIII (psoroftalmía), LXXIX
(xeroftalmía).
-
Nollete JA (1748). Quoted by McCaig et al. in: Controlling
Cell Behavior Electrically: Current Views and Future Potential.
-
Dutrochet H. Nouvelles recherches sur lendosmose et
l'exosmose. Paris :JB Ballière, 1828.
-
van Mohl., 1836. Quoted by Glasstone S. Textbook of Physical
Chemistry, Londres: Mac Millan, 1962.
-
Pfeffer W. Osmotische Untersuchungen. Leipzig. 1877. Quoted by
Glasstone S. Textbook of Physical Chemistry, Londres: Mac Millan, 1962.
-
Hartley GS. Trans. Faraday Soc 1937; 33: 1022.
-
Raoult FM. Loi de congélation des solutions aqueuses de
substances organiques. CR Acad Sci Paris 1882; 94:1517-1519.
-
MosesRA. Intraocular Pressure. Adlers Physiology of the Eye.
St.Louis:Mosby. 8ª edición. 1981. cap. VIII.
-
Ecoffet M, Demailly P, Koper J. Physiologie de lhumeur
aqueuse et de la tension oculaire. Encycl Med-Chir 1985. 21020 D10 (11-1985).
-
Cole DF. Aqueous and ciliary body. In :Graymore CN (ed.).
Biochemistry of the Eye. Nueva York :Academic Press, Inc. 1970.
-
Schaeffer AJ. Osmotic pressure of extraocular and intraocular
fluids. Arch Ophthalmol 1950; 43: 1026-1035.
-
Brubaker RF, Kupfer C. Mycroscopic determination of the
osmolality of interstitial fluid in the living rabbit cornea. Invest
Ophthalmol 1962; 1: 653-660.
-
Ridley F. The intraocular pressure and drainage of the aqueous
humor. Brit J Exp Path 1930; 11: 217-240.
-
Cogan DG, Kinsey VE. The cornea- Physiological aspects. Arch
Ophthalmol 1942; 28: 661-669.
-
Waltman SR. The Cornea. In Moses RA (ed.). Adlers Physiology
of the Eye. St.Louis:Mosby. 8ª edición. 1981. Cap. III.
-
Charlton CF. The function of the protein in the lacrimal
secretion. Am J Ophthalmol 1920; 3: 802-804.
-
von Bahr G . Könnte der Flüssigkeitsabgang durch die Cornea
von physiologischer Bedeutung sein? Acta Ophthalmol 1941; 19: 125-34.
-
Kinsey VE, Cogan DG. Arch Ophthalmol (Chicago) 1942; 28: 449
Citado por Ashton et al. Mechanism of corneal vascularization. Brit J
Ophthalmol 1953; 37: 193-209.
-
Giardini A, Roberts JRE. Conc entration of glucoe and total
chloride in tears. Brit J Ophthalmol 1950; 34: 737-743.
-
von Bahr G. Corneal thickness. Its measurement and changes. Am
J Ophthalmol 1956; 42: 251-263.
-
von Bahr G. The corneal thickness during perfusion of the
anterior chamber with solutions of various tonicity. Doc Ophthalmol 1962; 16:
3-10.
-
Yasuda H, Stone W. Theoretical study of the fluid transport
and the hydration of the cornea. J Theoret Biol 1957; 16: 111-134.
-
Scherz W, Doane MG, Dohlman CH. Tear volume in normal eyes and
keratoconjunctivitis sicca. Graefes Arch Ophthalmol 1974; 192: 141-150.
-
Michel J. Lehrbuch der Augenheilkunde. Wiesbaden. 1884. 1st
edition, p 2000. Citado por Schirmer O. Studien zur Physiologie und Pathologie
der Tränenabsorderung und Tränenabfuhr. Graefes Arch Ophthalmol 1903; 56:
197-291. p 242.
-
Massart J. Sensibilité et adaptation des organismes a la
concentration des solutions salines. Arch Biol 1889; 9: 515-570.
-
Mishima S. Some physiological aspects of the precorneal tear
film. Arch Ophthalmol 1965; 73: 233-241. Quoted by Francisci et al 1980.
-
Magaard H. Ueber das Secret und die Secretion der menschlichen
Thänendrüse. Virchows Arch pathol Anat 1882; 89: 258-271.
-
Beaunis H. Nouveaux éléments de physiologie humaine. 3rd
edition, Paris. 1888.
-
Hambürger HJ. Osmotischer Druck und Ionenlehre. 1904;III. 162
. Quoted by Cantonnet 1908a. como de 1906 y por Pedersen-Bjergaard (más
creible) como de 1904.
-
Cantonnet A. Solution chlorurée isotonique aux larmes pour
lavages et bains de loeil. (Saline solution isotonic with the tears as an eye
wash). Arch Ophtalmol (Fr). 1908.by Cantonnet 1908b. Abstract in Arc Oftalmol
Hiso-Amer 1909; 9: 385.
-
Cantonnet A. Formules de collyres isotoniques aux larmes.
(Formulae of collyria isotonic with the tears) Arch Ophtalmol (Paris) 1908;
28: 617-621.
-
von Rötth A (De Roetth A). Ueber die Tränenflüssigkeit. Klin
Monatsbl Augenheilk 1922; 68: 598-604.
-
Michaïl D. Recherches sur lélimination lacrymale du chlorure
de sodium. Ann dOcul (Paris) 1936; 173: 715-734.
-
Krogh A, Lund CG, Pedersen-Bjergaard K. The osmotic
concentration of human lachrymal fluid. Acta Physiol Scand 1945; 10: 88-90.
-
Schaeffer AJ. Osmotic pressure of extraocular and intraocular
fluids. Arch Ophthalmol 1950; 43: 1026-1035.
-
Janot MM. Nouveaus aspects scientifiques de la Pharmacopée.
Annales Pharmac Franç 1951; 9: 129-140.
-
Balik J. The lacrimal fluid in keratoconjunctivitis sicca: A
quantitative and qualitative investigation. Am J Ophthalmol 1952; 35: 773-782.
-
Pedersen-Bjergaard K, Smidt BC. Electrolytic conductivity,
osmotic pressure and hydrogen ion concentration of human lachrymal fluid. Acta
Dermat Venereol. 1952. suppl 29; 31: 265-267.
-
Potts AM. The nutritional supply of corneal regions in
experimental animals. II. The problem of corneal transparency and the tonicity
of tears. Am J Ophthalmol 1953; 36: 127-138.
-
Mastman GJ, Baldes EJ, Henderson JW. The total osmotic
pressure of tears in normal and various pathologic conditions. Arch Ophthalmol
1961; l 65: 509-513.
-
Mishima S, Gasset A, Klyce SD, Baum JL. Determination of tear
volume and tear flow. Inv Ophthalmol Vis Sci 1966; 5: 264-276.
-
Mishima S. Corneal thickness. Surv Ophthalmol 1968; 13: 57-96.
-
Mishima S, Kubota Z, Farris RL.The tear flow dynamics in
normal and in keratoconjunctivitis sicca cases. XXI Concilium Ophthalmologicum
(Mexico 1970). Amsterdam: Excerpta Medica, 1971. Pars II, pp 1801-5.
-
Milder B. The lacrimal apparatus In: Moses RA (ed). Adlers
physiology of the eye; clinical application. 6th edition Saint Louis:Ed.
Mosby. 1975. Pp 17-34.
-
Métaireau JP, Baikoff G, Brun P. La physiologie lacrymale.
Arch Ophtalmol (Paris) 1977 ;37: 401-408.
-
Terry JE, Hill RM. Osmotic adaptation to rigid contact lenses.
Arch Ophtalmol (Paris) 1977; 37: 785-788.
-
Terry JE, Hill RM. Human tear osmotic pressure. Diurnal
variations and the closed eye. Arch Ophthalmol 1978; 96: 120-122.
-
Gilbard JP, Farris RL, Santamaría J. Osmolarity of tear
microvolumes in keratoconjunctivitis sicca. Arch Ophthalmol 1978; 96: 677-681.
-
Benjamin WJ, Hill RM. Human tears : osmotic characteristics.
Invest Ophthalmol Vis Sci 1983; 24: 1624-1626.
-
Gilbard JP, Gray KL, Rossi SR. A proposed mechanism for
increased tear-film osmolarity in contact lens wearers. Am J Ophthalmol 1986;
102-505-7.
-
Nelson JD, Wright JC. Tear film osmolality determination: an
evaluation of potential errors in measurement. Curr Eye Res 1986; 5: 677-681.
-
Murube J, Cortés Rodrigo MD. Eye parameters for the diagnosis
of xerophthalmos. Clin Exp Rheumatol 1989; 7: 141-150.
-
Lucca JA, Nunez JN, Farris RL. A comparison of diagnostic
tests for keratoconjunctivis sicca: lactoplate, Schirmer, and tear osmolarity.
CLAO J 1990; 16: 109-112.
-
Sullá i Molina O, Quintana i Conte R, Sempere MA, Losada M.
Determinación de la osmolaridad lagrimal en el síndrome de ojo seco. Arch Soc
Españ Oftalmol 1992; 62: 503-506.
-
Gilbard JP, Rossi SR. Changes in tear ion concentrations in
dry-eye disorders. Adv Exp Med Biol 1994; 350: 529-533.
-
Sullivan BD, Angeles R, Lemp MA. Clinical results of a first
generation lab-on-a-chip nanoliter tear film osmometer. The Ocular Surface
2005; 3(suppl): S117.
-
Lemp MA. A new nanoliter tear osmometer for the clinical
diagnosis of dry eye. Abstracts of the 8th Congress of the International
Society of Dacryology and Dry Eye. Madrid 2005
-
Gilbard JP, Farris RL. Tear osmolarity and ocular surface
disease in keratoconjunctivitis sicca. Arch Ophthalmol 1979; 97: 1642-1646.
-
Gilbard JP, Kenyon KR . Tears diluents in the treatment of
keratoconjunctivitis sicca. Ophthalmology 1985; 92: 646-650.
-
Lucca JA, Farris RL, Bielory L, et al. Keratoconjunctivitis
sicca in male patients with human immunodeficiency virus type I.
Ophthalmology, 1990; 97: 1008-10.
-
Sust Mir F. Farmacotecnia oftalmológica. Arch Soc Oftalmol
Hisp-SAmer 1967; 27: 313-38.
-
Holly FJ, Lamberts DW. Effect of nonisotonic solutions on tear
osmolarity of tear substitutes. Inv Ophthalmol Vis Sci 1981; 20: 236-245.
-
Cantonnet A. Causeries dOphtalmologie. Paris: Maloine. 1925.
p 219-20 and 290-1.
-
Maurice DM. The tonicity of an eye drop and its dilution by
tears. Exp Eye Res 1971; 11: 30-3.
-
Egbert PR, Lauber S, Maurice DM. A simple conjunctival biopsy.
Am J Ophthalmo 1977; 84: 798-801.
-
Huang AJ, Tseng SC,Kenyon KR. Paracellular permeability of
corneal and conjunctival epithelia. Invest Ophthalmol Vis Sci 1989; 30:
684-689.
-
Vergés CM, Gilbard JP, Refojo MF, et al. Effect of
hyperosmolarity on conjunctival goblet cells density in vivo. Inv Ophthalmol
Vis Sci 1984; 25: 191.
-
Vergés Roger C, Pita Salorio D . Efecto de la hiperosmolaridad
sobre la población de las células caliciformes en la conjuntiva y la secreción
de mucina. Arch Soc Españ Oftalmol 1985; 48: 293-298.
-
Vergés Roger C, Pita Salorio D, Refojo MF, Sainz de la Maza
Serra MT. Cambios en la población de células caliciformes conjuntivales tras
la aplicación de una solución hiperosmolar. Arch Soc Españ Oftalmol 1986; 51:
403-406.
-
Vergés Roger C, Pita Salorio D. Estudio de la población de
células caliciformes conjuntivales en los síndromes de ojo seco mediante un
método citológico atraumático. Arch Soc Españ Oftalmol 1988; 55: 525-530.